МЕТАБОЛИЗМА
Зрелые красные кровяные тельца обладают двояковогнутой формой и большой способностью к деформации, благодаря чему эффективно обеспечивают процессы диффузии газов и могут проходить через капилляры, диаметр которых в 3-4 раза меньше самих эритроцитов. Подобное свойство обусловлено особенностями в структуре мембран этих форменных элементов.
Принципы строения цитолеммы эритроцитов классические: основу составляет билипидный слой, в который встроены различные протеины. Наружная часть липидов представлена холинсодержащими соединениями (фосфатидилхолином, сфингомиелином), внутренняя поверхность обогащена фосфатидилсерином, фосфатидилэтаноламином, более четверти объёма приходится на холестерин. К цитозолю обращена плотная анастомозирующая белковая сеть, состоящая из спектринов, анкиринов, тропомиозинов, которые связываясь с интегральными гликопротеидами (гликофоринами) создают определённую жёсткость мембраны, определяют форму эритроцита (двояковогнутый диск). От степени фосфорилирования спектринов зависит эластичность сети, способность к упругой деформации. Гликофорины, пронизывая липидный слой, с помощью гидрофобных взаимодействий с фосфолипидами прочно фиксируются; их углеводные компоненты (олигосахариды, их производные, сиаловые кислоты), располагаясь на поверхности мембраны эритроцита, служат групповыми веществами крови: для О(Н) антигена детерминанта фукоза, для А-антигена - N-ацетилгалактоз-амин, для В – галактоза. Углеводные остатки гликофоринов являются также носителями антигенов групп крови MN-типа и ряда других иммунологических детерминант, служат рецепторами для вирусов гриппа и т.д.
Несмотря на отсутствие ядра и других органоидов, красные кровяные тельца весьма метаболически активные образования, обладающие спонтанной энергопродукцией на уровне 80 пДж на клетку. Энергетика эритроцита основана на анаэробном гликолизе (рис. 1). Глюкоза быстро проникает в клетку, независимо от присутствия инсулина.
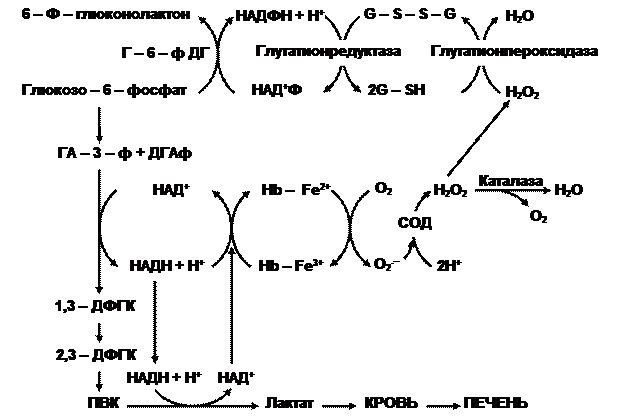
Рис.1. Схема гликолиза и антирадикальной защиты в эритроците.
В пути Эмбдена-Мейергофа глюкоза распадается до лактата с образованием АТФ путём субстратного фосфорилирования. Следует заметить, что кроме энергетической функции, метаболиты гликолиза используются в следующих процессах. Восстановительные потенциалы генерируются в виде НАДН, который при необходимости используется метгемоглобин-редуктазой для восстановления железа в метгемоглобине. В отличие от других тканей в эритроцитах в качестве метаболита образуется много 2,3-дифосфоглицерата (2,3-ДФГК) (в 1000 раз больше), который служит важным модулятором сродства гемоглобина к кислороду.
Некоторые морфобиохимические особенности красных кровяных телец предопределяют необходимость в высокой антиоксидантной активности. Во-первых, это высокие концентрации О2, что увеличивает вероятность образования его активных форм. Во-вторых, большое содержание ионов переходного металла – железа, что может способствовать его использованию в качестве донора электронов (рис.1). И, наконец, для обеспечения упругой деформации в липидном бислое мембран содержится больше ПНЖК – субстратов ПОЛ, чем в цитолемме других клеток. Для контроля интенсивности свободнорадикальных процессов, в цитоплазме эритроцитов активно работает антирадикальная защита.
Если нарушаются условия диссоциации оксигемоглобина (в кармане глобина, где расположен гем, оказываются ионы, молекулы воды), то в этом случае происходит отрыв электрона от двухвалентного железа гема с образованием метгемоглобина и супероксидного анион-радикала. Первое соединение восстанавливается с помощью метгемоглобинредуктазы, а радикал кислорода преобразуется под влиянием супероксиддисмутазы(СОД) в пероксид водорода, который также токсичен для клеток. Поэтому он восстанавливается с первоначально помощью каталазы, позднее глутатионпероксидазы (ГПО) (рис.1) и восстановленного глутатиона. Чтобы поддержать пул последнего, работает глутатионредуктаза (ГР), которая восстанавливает окисленную форму пептида, применяя в качестве восстановителя НАДФН. Необходимую концентрацию этого варианта кофермента получают путем окисления глюкозо-6-фосфата соответствующей дегидрогеназой. В мембранах эритроцитов продуктами свободнорадикальных процессов могут быть липопероксиды (ROOH), которые своевременно удаляются восстановленным глутатионом с участием ГПО:
| 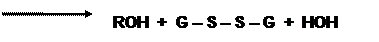 |
Дата добавления: 2015-06-05; просмотров: 1390;