Т-лимфопоэз в тимусе.
Предшественники Т-лимфоцитов, покидая костный мозг, через кровь мигрируют в тимус, которого достигают не более 5% клеток. Часть "затерявшихся клеток", попадая в селезенку, может дифференцироваться в NK- клетки. Миграция предшественников Т-лимфоцитов в тимус осуществляется
благодаря их высокой подвижности, наличию на поверхности клеток рецепторов хоминга (CD44 - проводник клеток в тимус), а также действию хемокинов. Развитие Т-лимфоцитов в тимусе обеспечивается взаимодействием с тимическими эпителиальными клетками (ТЭК).
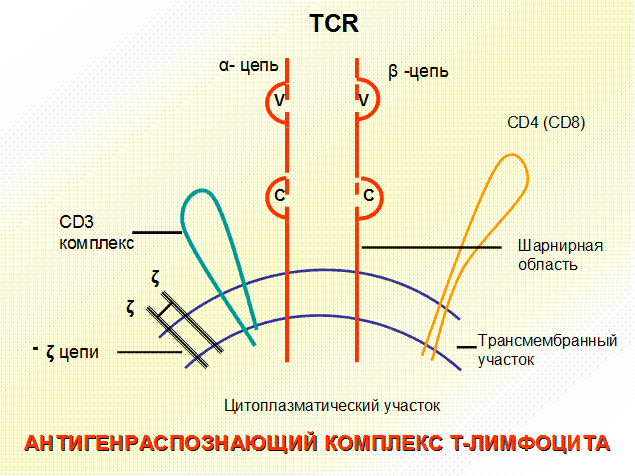
Рис. 18. Антигенраспознающий комплекс Т-лимфоцита
ТЭКсинтезируют гормоны тимуса (тимулин, тимозины, тимопоэтин, сывороточный фактор тимуса), которые стимулируют дифференцировку и функциональную активность Т-лимфоцитов. Стромальные клетки тимуса продуцируют и секретируют целый ряд ростовых факторов и других цитокинов: ИЛ-1, 6, 7, 11, 12, ФНО-α, ТФР-b, GM-CSF и др. Важнейшим для интратимической дифференцировки Т-лимфоцитов является ИЛ-7. В процессе ИЛ-7-опосредованной пролиферации в субкортикальной зоне формируются незрелые Т-клетки, экспрессирующие CD25.Клетки с фенотипом CD44+CD25+обозначаются как проТ,на этой стадии начинается реаранжировка рецепторных генов. В субкортикальном слое начинает экспрессироваться CD3 рецептор, состоящий из γ, β и έцепей, и обеспечивающий передачу от сигнала TCR в цитоплазму клетки.
Кроме того, ранние предшественники Т-лимфоцитов несут на своей поверхности CD34, CD38 (маркер стадии синтеза), CD7, TdT (терминальная дезоксинуклеотидил трансфераза - исчезает по мере созревания клеток; необходима для встраивания дополнительных нуклеотидных последовательностей в ДНК клетки, что увеличивает структурное разнообразие рецепторов).
На начальном периоде пребывания в тимусе клетки способны дифференцироваться в направлении Т- лимфоцитов,В-,NK - , дендритных клеток. Эта способность утрачивается после потери CD44.
На стадии пре-Т -клеток (CD44-CD25-)завершается реаранжировка рецепторных генов. На этой стадии происходит выбор в пользу формирования рецепторов α/β или γ/δ. Одновременно с этим (кортикальный слой) усиливается экспрессия CD4+ (обеспечивает распознавание аг ГКГС 2 класса)и CD8+ (обеспечивает распознавание аг ГКГС 1 класса)(CD4+CD8+ - дубль-положительные клетки).Для незрелых (кортикальных) тимоцитов также характерны CD1 (a, b, c), CD95(проводник в клетку сигнала апоптоза). Кортикальные тимоциты очень чувствительны к действию индукторов апоптоза. Зрелые медуллярные тимоциты экспрессируют CD28+ (на 60-70% СD8+, на всех CD4+ - корецептор межклеточного взаимодействия, предохраняет клетки от апоптоза), CD45 (участвует в запуске активационных сигналов).
В процессе созревания тимоцитов изменяется их фенотип, свойства, происходит селекция Т-лимфоцитов, основные процессы осуществляются на границе коркового и мозгового слоев.
Этапы селекции Т – лимфоцитов
1. Образование дубль положительных клеток.
2. Отбор клонов клеток, способных распознавать собственные молекулы ГКГС 1 или 2 класса, экспрессированные на ТЭК (тимоциты, не способные распознать собственные антигены ГКГС гибнут путем апоптоза).
3. Клетки, распознавшие молекулу ГКГС 1 класса теряют CD4, сохраняют CD8; клетки, распознавшие молекулу ГКГС 2 класса теряют CD8, сохраняют CD4 (положительная селекция).
4. Отрицательная селекция: из популяции тимоцитов через процесс апоптоза удаляются клетки, способные реагировать с аутоантигенами на поверхности АПК.
5. Оставшиеся от исходной популяции клеток 2-5% покидают тимус и заселяют периферические лимфоидные органы и ткани.
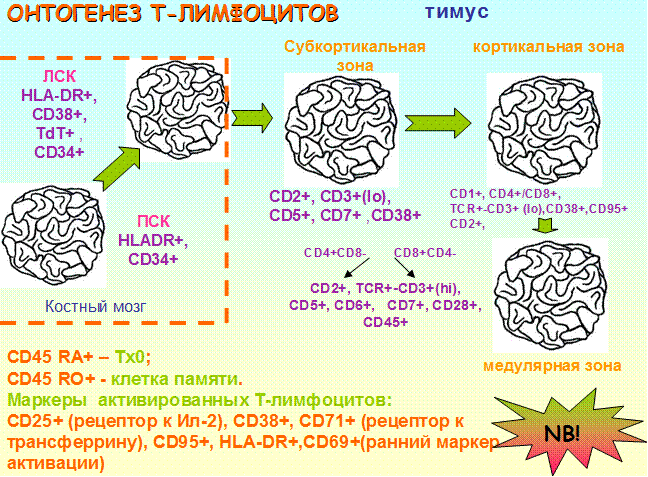
Рис. 19. Онтогенез Т-лимфоцитов
Т-лимфоциты – продуценты цитокинов
| Клетки-продуценты | Спектр цитокинов |
| Тх0 | ИЛ-2, 3,4,5,6, 10,13; ИФН-g, ФНО-b |
| Тх1 | ИЛ-2, 3, ИФН-g, ФНО-b |
| Тх2 | ИЛ-3,4,5,6, 10,13 |
| Т reg | ИЛ – 5, 10, ИФН-g, ТФР-b |
| ЦТЛ | ИЛ-2,4,5,10, ИФН-g, ФНО-g |
Дата добавления: 2015-06-05; просмотров: 3084;