ЧУВСТВИТЕЛЬНЫЕ РЕЦЕПТОРЫ, СВЯЗАННЫЕ С РАСТЯГИВАНИЕМ
Три вида рецепторов непосредственно связаны с растягиванием и поддержанием оптимального диапазона движения. Это нервно-мышечные веретена, нервно-сухожильные веретена и суставные механорецепто-ры. Далее мы рассмотрим их структуру, функции и взаимосвязь с растягиванием.
Нервно-мышечные веретена. Основными тензорецепторами мышцы являются нервно-мышечные веретена. Это наиболее подробно изученные проприорецепторы — рецепторы, расположенные в мышцах, сухожилиях и преддверии уха. Их рефлексы связаны с локомоцией или положением тела. В разном количестве их можно обнаружить в большинстве скелетных мышц тела. Особенно много нервно-мышечных веретен содержится в небольших мышцах кисти и глаза. Поскольку мышечные
Наука о гибкости
 волокна заключены в веретенообразную капсулу (оболочку соединительной ткани, окутывающую рецептор), их называют интрафузальными волокнами. Экстрафузальные волокна являются обычными сократительными единицами мышцы. Веретена прикрепляются к обоим концам экстра-фузальных волокон и, таким образом, располагаются параллельно им. Следовательно, при растягивании мышцы веретено также подвергается растягиванию.
волокна заключены в веретенообразную капсулу (оболочку соединительной ткани, окутывающую рецептор), их называют интрафузальными волокнами. Экстрафузальные волокна являются обычными сократительными единицами мышцы. Веретена прикрепляются к обоим концам экстра-фузальных волокон и, таким образом, располагаются параллельно им. Следовательно, при растягивании мышцы веретено также подвергается растягиванию.
Веретено интрафузальныхмышечных волокон
Существует два основных типа интрафузальных волокон: волокна ядерного мешочка и волокно ядерной цепочки. Первые содержат в большом количестве саркоплазму и клеточные ядра в расширенной, напоминающей мешок структуре. Эта несократительная структура находится в центральном, или экваториальном, участке интрафузального волокна. Отсюда и название — интрафузальное волокно ядерного мешочка. На дистальных, или полярных, концах волокна ядерного мешочка находятся исчерченные сократительные филаменты, которые возле веретен прикрепляются к эк-страфузальным волокнам. Не так давно было установлено, что существует два подвида волокон ядерного мешочка. Их назвали мешочек1 и мешоче^ (GJadden, 1986; рис. 6.5).
Второй тип интрафузального мышечного волокна — волокно ядерной цепочки — более тонкое и короткое, чем волокно ядерного мешочка. Оно содержит всего один ряд ядер, расположенных в виде цепочки по несократительному экваториальному участку. Подобно волокнам ядерного мешоч-
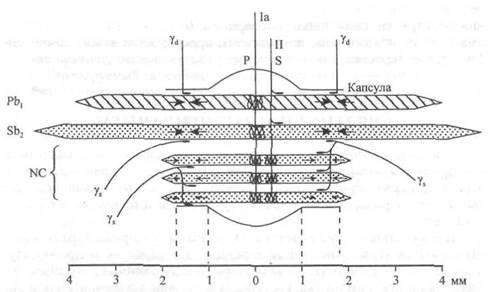
Рис 6.5. Взаимосвязь первичного (Р) и вторичного (S) чувствительных нервных окончаний, иннервируемых аксонами группы 1а и И, соответственно с тремя видами интрафузальных волокон, мешочек, (Ь,), мешочек, (Ь,) и волокнами ядерной цепочки (NC) и их двигательными окончаниями (Gladden, 1986) ~ '
Глава 6. Нейрофизиология гибкости: невральная анатомия и физиология
 ка, полярные концы волокна ядерной цепочки также состоят из исчерченных сократительных филаментов. Их концы нередко соединяются с волокнами ядерного мешочка, которые, в свою очередь, прикрепляются к эндо-мизию экстрафузальных волокон.
ка, полярные концы волокна ядерной цепочки также состоят из исчерченных сократительных филаментов. Их концы нередко соединяются с волокнами ядерного мешочка, которые, в свою очередь, прикрепляются к эндо-мизию экстрафузальных волокон.
Чувствительные нервы веретен
Существует два вида чувствительных нервных (афферентных) окончаний в каждом веретене: первичные и вторичные. Первичные окончания заканчиваются, спиралевидно окружая центральный участок волокна ядерного мешочка и выпуская боковое ответвление к волокну ядерной цепочки. Афферентные аксоны первичных нервных окончаний относятся к волокнам большой группы I. Чтобы отличить эти чувствительные нервные окончания от других, относящихся к этой же группе, большие афференты веретена называют афферентами группы 1а.
Первичные нервные окончания имеют очень низкий порог растяжения и поэтому легко возбуждаются. Их реакции могут быть как фазными (динамическими), так и тоническими. Фазная реакция определяет интенсивность или скорость растягивания путем изменения частоты нейронных импульсов во время растягивания. Частота разрядки быстро увеличивается при начальном растягивании. Затем, по достижении новой длины, она снижается до постоянного уровня, соответствующего новой тонической длине. Следовательно, тоническая реакция определяет длину мышцы. Иными словами, первичные нервные окончания определяют длину и скорость растягивания.
Вторичные нервные окончания образуют разветвленные, напоминающие цветок окончания. Они содержатся в основном в околоэкваториальном сегменте (вблизи экватора) волокон ядерной цепочки. Аксоны вторичных нервных окончаний относятся к афферентным волокнам группы II. В отличие от первичных нервных окончаний, вторичные нервные окончания определяют только тоническую длину мышцы.
Эти мотонейроны вызывают сокращение мышечных филаментов в полярных концах интрафузальных мышечных волокон. При сокращении этих концов в центральном экваториальном участке наблюдается пассивное растягивание (там находятся чувствительные нейронные рецепторы). Таким образом, активация гамма-мотонейронов ЦНС может увеличивать количество растяжения, испытываемого чувствительными нервными окончаниями (Banker, 1980).
Двигательные нервы веретена
Двигательные нервы, иннервирующие каждое интрафу зальное мышечное волокно в его полярных участках, называются у-эфферентны-ми волокнами, образующими фузимоторную систему. Различают два вида у-аксонов в зависимости от их воздействия на первичные и вторичные чувствительные нервные окончания- Стимулирование так называемого статического 7-аксона (ys) может усиливать чувствительность первично-
Наука о гибкости
го нервного окончания к длине, не влияя (или незначительно влияя) на восприятие (чувствительность) скорости. С другой стороны, динамические у-аксоны (у.) могут существенно усиливать восприятие скорости первичного нервного окончания, практически не влияя на чувствительность к длине.
Функция у-системы заключается в контроле чувствительности веретена к растягиванию. Этот процесс сенситизации веретена при помощи у-эф-ферентов известен как у-смешение (Norback и Demarest, 1981). Активация у-мотонейронов приводит к сокращению или укорачиванию интрафузаль-ных мышечных волокон в их полярных участках. При сокращении интра-фузальных волокон экваториальный участок растягивается. Эта центральная деформация приводит к перетягиванию колец в аннулоспиральную часть первичных окончаний и увеличению интенсивности разрядки аффе-рентов групп 1а и II.
Вторая функция 7-мотонейронов — поддержание чувствительности веретена во время сокращений мышцы. Когда мышцы укорачиваются, веретено также пассивно укорачивается. Это пассивное сближение двух концов веретена снимает напряжение как с его первичных окончаний (разгрузка веретена), так и со вторичных; такая разгрузка лишает головной мозг информации из веретена относительно изменений мышечной длины. Чтобы предотвратить разгрузку веретена и отрегулировать его чувствительность, у-мотонейроны активируют.
Процесс возбуждения нервно-мышечного веретена
Процесс возбуждения нервно-мышечного веретена можно охарактеризовать следующим образом. Во-первых, к нервно-мышечному веретену прикладывают минимальный растягивающий стимул. Во-вторых, в чувствительном нейронном окончании происходит изменение степени проницаемости, что приводит к производству генерирующего тока (передача заряда через мембрану нервного терминала). Последний, в свою очередь, вызывает деполяризацию, которая называется генератор потенциала. При несколько большей степени растягивания его амплитуда в окончаниях нервно-мышечного веретена увеличивается. Когда деполяризация достигает порога, возникает проведенный потенциал действия. При дальнейшем увеличении степени растягивания может наблюдаться серия проводимых нервных импульсов. Этапы возбуждения нервно-мышечного веретена приведены ниже:
 (О Р) (3( (4) (5)
(О Р) (3( (4) (5)
 Стимул Локальное Генератор Локальная де поляр и- Проведенный
Стимул Локальное Генератор Локальная де поляр и- Проведенный
(растягивание) изменение тока (передача зация (генератор потенциал
проницаемости заряда) потен циала) действия
Нервно-сухожильные веретена. Нервно-сухожильные веретена — это чувствительные к сокращению механорецепторы скелетных мышц
Глава 6. Нейрофизиология гибкости: неврааьная анатомия и физиология
 млекопитающих, иннервируемые быстропроводящими афферентными нервными волокнами группы 16, имеющими большой диаметр (Jami, 1992). Впервые нервно-сухожильные веретена были обнаружены и описаны Гольджи в 1903 году. Ввиду технических трудностей нервно-сухожильные веретена не так хорошо изучены, как нервно-мышечные. Тем не менее их важность с точки зрения развития гибкости нельзя недооценивать.
млекопитающих, иннервируемые быстропроводящими афферентными нервными волокнами группы 16, имеющими большой диаметр (Jami, 1992). Впервые нервно-сухожильные веретена были обнаружены и описаны Гольджи в 1903 году. Ввиду технических трудностей нервно-сухожильные веретена не так хорошо изучены, как нервно-мышечные. Тем не менее их важность с точки зрения развития гибкости нельзя недооценивать.
Местонахождение и структура нервно-сухожильных веретен
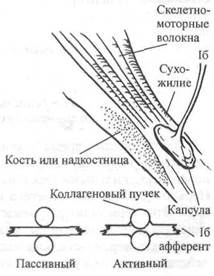
Нервно-сухожильные веретена находятся почти исключительно у апоневрозов или мышечно-сухожильных соединений, но не в сухожилиях (рис. 6.6). Так, Пенг, исследовав (цитируется по Bavker, 1974) 1337 рецепторов из различных участков мыши кошек, установил, что 92,4 % их располагались у мышечно-сухожильных соединений и только 7,6 % — в самом сухожилии.
|
У немлекопитающих нервно-сухожильные веретена представляют собой расположенные вдоль сухожильных пучков не покрытые оболочкой рецепторы (пучки коллагеновых волокон). У млекопитающих же, напротив, они покрыты оболочкой и, как уже упоминалось, располагаются у мышечно-сухожильных или мышечно-апоневротических соединений. Значение этого различия, по мнению исследователей, состоит в следующем. Во-первых, окруженные оболочкой веретена являются более чувствительными к данной величине стимула и более точными в определении и передаче информации в ЦНС. Во-вторых, местонахождение нервно-сухожильных веретен обеспечивает их экстремальную чувствительность к любому изменению напряжения отдельных мышечных волокон, к которым они прикреплены (J.C.Moore, 1984).
| Рис. 6.6. Сухожильный аппарат Голь джи. Покрытый оболочкой афферент 16 окружает сухожилия нескольких скелет но-моторных волокон. Сокраще ние этих волокон вызывает сжатие кол лагеновых пучков (сухожилие) и, та ким образом, активирует афферент 16 . (Enoka, I988) |
Нервно-сухожильные веретена располагаются последовательно, в отличие от нервно-мышечных веретен, расположенных параллельно мышечным волокнам. В исследовании Пенга количество мышечных волокон, соединенных последовательно с нервно-сухожильными веретенами, колеблется от 3 до 50. Каждое нервно-сухожильное веретено, как правило, иннервируется отдельным быстро проводящим афферентным нервным волокном группы 16 (Jami, 1992).
Наука о гибкости
 Фунтсцш нервно-сухожильных веретен
Фунтсцш нервно-сухожильных веретен
На данном этапе мы лишь частично знаем многочисленные функции нервно-сухожильных веретен. Последние исследования показывают, что эти рецепторы являются более сложными, чем первоначально считалось (J.C.Moore, 19S4). Сейчас мы знаем, что нервно-сухожильные веретена контролируют все степени мышечного напряжения. В то же время они более всего чувствительны к напряжению, производимому в результате мышечных сокращений (см. рис. 6.6). Предполагают также, что нервно-сухожильные веретена вносят свой вклад в сознательные восприятия. Это предположение основано на том факте, что импульс из нервно-сухожильных веретен достигает коры головного мозга, той его части, которая осуществляет интерпретацию сенсорной активности тела (Roland и Ladegaard-Pedersen, 1977).
Кроме того, высказывается мнение, что нервно-сухожильные веретена способствуют снижению мышечного утомления (Вагг, 1979; Kandel, Swartz, 1981; Lundberg, 1975). По мере увеличения напряжения в группе мышечных волокон нервно-сухожильные веретена и их афференты группы 16 посылают увеличивающееся число сигналов в ЦНС. Эти чувствительные нервы заканчиваются в спинном мозгу на небольших интерней-ронах, которые затем ингибируют клеточные тела мотонейронов, активирующих сокращающуюся мышцу. Данный процесс называется автогенным ингибированием, так как сокращения мышцы ингибируются ее собственными рецепторами. Результирующее снижение мышечной силы уменьшает активацию нервно-сухожильных веретен и количество инги-биторной обратной связи, поступюшей в ЦНС из этих мышц. Такое мгновенное ингибирование обратной связи способствует увеличению напряжения в мышцах.
Неправильные представления
о нервно-сухожильных веретенах
За прошедшее время появилось немало ошибочных представлений о нервно-сухожильных веретенах. Одно из них касается взаимосвязи между нервно-сухожильными веретенами и измерением растягивающих усилий. Нервно-сухожильные веретена нередко называют «рецепторами растяжения». Этот термин подразумевает, что пассивное напряжение, сопровождающее растягивание мышцы, может также представлять собой адекватный стимул для нервно-сухожильных веретен. Нервно-сухожильные веретена, действительно, могут активироваться в результате пассивного напряжения, однако их порог для данного вида стимулов является очень высоким. Поэтому для активизации нервно-сухожильных веретен необходимо очень интенсивное растягивание (Houk, Singer, Goldman, 1971). Кроме того, разрядка нервно-сухожильных веретен очень редко продолжается во время поддерживаемого растяжения мышцы.
Другое неверное представление относительно нервно-сухожильных веретен заключается в том, что их чувствительность является недостаточ-
Глава 6. Нейрофизиология гибкости: неврапъная анатомия и физиология
 ной для активизации сократительного напряжения. Исследования показали, что нервно-сухожильные веретена проявляют очень низкий порог и весьма ощутимую динамическую чувствительность при тестировании с применением адекватного стимула. Таким образом, нервно-сухожильные веретена способны сообщать об очень незначительных и быстрых изменениях в силах сокращения (Houk и Henneman, 1967; Houk, Singer, Goldman, 1971; Jami, 1992).
ной для активизации сократительного напряжения. Исследования показали, что нервно-сухожильные веретена проявляют очень низкий порог и весьма ощутимую динамическую чувствительность при тестировании с применением адекватного стимула. Таким образом, нервно-сухожильные веретена способны сообщать об очень незначительных и быстрых изменениях в силах сокращения (Houk и Henneman, 1967; Houk, Singer, Goldman, 1971; Jami, 1992).
Некоторое время существовало мнение, что функция нервно-сухожильных веретен заключается в автогенном ингибировании, т. е. ингиби-ровании агонистов и синергистов и содействии антагонистам. В настоящее время доказано, что это лишь одна из многих функций нервно-сухожильных веретен, которым помогают в этом, по меньшей мере, еще два рецептора: низкопороговые рецепторы суставной капсулы и низкопороговые кожные рецепторы (J.С.Moore, 1984). Цель такого аутогенного ингибиро-вания связывают с защитными функциями.
Таблица 6.1. Характеристика системы суставных рецепторов
 Тип
Тип
Морфология
Местонахождение
Источник нервных волокон
Бн хев и орал ь ная характеристика
| I | Шарообразные частицы | Фиброзные капсулы | Небольшой | Статические и |
| (100 мкм - 40 мкм), | суставов | миелиниро- | динамические | |
| покрытые тонкой | (в поверхностных | ванный | механорецепторы: | |
| оболочкой в трехмерных | слоях) | (6-9 мкм) | низкий порог, | |
| кластерах, состоящих из | медленноадапти- | |||
| 3-8 частиц | рующиеся | |||
| II | Конические частицы | Фиброзные капсулы | Средний | Динамические |
| (280 мкм ■ 120 мкм), | суставов (в глубо- | миелиниро- | механорецепторы: | |
| покрытые толстой обо- | ких субсиновиаль- | ваниый | низкий порог, | |
| лочкой, отдельные или в | ных слоях). Сустав- | (9-12 мкм) | быстроадапти- | |
| кластерах по 2—4 частицы | ные жировые тела | рующиеся | ||
| III | Веретенообразные части- | Поверхность | Большой | Динамические |
| цы (600 мкм ■ 100 мкм), | суставных связок | миелиниро- | механорецепторы: | |
| покрытые тонкой оболоч- | (коллатеральные | ванный | высокий порог. | |
| кой, отдельные или в | и внутренние) | (13-17 мкм) | медл ен ноадапти - | |
| кластерах по 2-3 | рующиеся | |||
| IV | (а) Трехмерные сплетения | Фиброзные капсулы | Очень | Болевые рецепто- |
| немиел и н и ро в анн ы X | суставов. Суставные | маленькие | ры: очень высокий | |
| нервных волокон | жировые тела. | миелиниро- | порог, неадапти- | |
| Адвентициальные | ванные | рую щиеся. Гемо- | ||
| оболочки | (2-5 мкм) и | чувств ител ь н ые | ||
| кровеносных | немиелини- | (к аномальным | ||
| сосудов | рованные | метаболитам | ||
| (< 2 мкм) | ткани) болевые | |||
| рецепторы | ||||
| (б) Свободные | Суставные связки | |||
| не м и ел и н ирован н ы е | (коллатеральные и | |||
| нервные окончания | внутренние) |
 Наука о гибкости
Наука о гибкости
 Также известно, что влиянию нервно-сухожильных веретен могут противостоять дополнительные сигналы, поступающие из высших центров. Процесс снижения к минимуму влияния нервно-сухожильных веретен называется растормаживайием мотонейронов-агонистов. О важности этого процесса для спорта говорили Брукс и Фехи (1987):
Также известно, что влиянию нервно-сухожильных веретен могут противостоять дополнительные сигналы, поступающие из высших центров. Процесс снижения к минимуму влияния нервно-сухожильных веретен называется растормаживайием мотонейронов-агонистов. О важности этого процесса для спорта говорили Брукс и Фехи (1987):
«Использование растормаживания применяется в спортивной тренировке с целью довести физическую работоспособность до пределов возможности тканей. В таком виде спорта, как армрестлинг, иногда встречаются разрывы мышц и сухожилий и переломы костей. У высокомотивированных спортсменов и у расторможенных индивидуумов сочетание активного мышечного сокращения и напряжения, производимого соперником, может превысить силу тканей».
Суставные механорецепторы (рецепторы суставов). Все синовиальные суставы тела имеют четыре различных нервных окончания. Эти суставные рецепторы ощущают механические силы, действующие на суставы, такие, как давление растяжения, и поэтому их называют суставными механо-рецепторами. Выделяют четыре типа суставных механорецепторов (табл. 6.1). Такая классификация основана на соответствующих морфологических и бихевиоральных характеристиках нервных окончаний. В следующих разделах мы рассмотрим эти четыре вида суставных механорецепторов.
Суставные рецепторы типа I
Механорецепторы типа I состоят из кластеров тонких, покрытых оболочкой шарообразных частиц. Онн находятся главным образом во внешних (поверхностных) слоях фиброзной суставной капсулы. Каждый кластер состоит приблизительно нз 8 частиц. Каждая частица иннервируется отдельным миелинированным волокном группы П (диаметром 6-9 мкм). Плотность этих механорецелторов более высокая в проксимальных (например, тазобедренный), чем в дистальных (например, голеностопный) суставах.
С физиологической точки зрения частицы типа I ведут себя как низкопороговые медленноадаптирующиеся рецепторы. Следовательно, они реагируют на очень незначительную механическую нагрузку и продолжают выпускать нервные импульсы на протяжении действия механического стимула. Чтобы стимулировать их, достаточно усилие порядка 3 г. Более того, часть этих рецепторов низкого порога всегда активна при любом положении сустава и даже в том случае, когда он неподвижен. Их интенсивность разрядки в покое обычно имеет частоту примерно 10-20 Гц (импульсов в секунду).
Механорецепторы типа I имеют целый ряд функций, включая сообще
ние направления, амплитуды и скорости движений суставов; регуляцию
изменений давления в суставе; содействие постуральным и кинестетичес
ким ощущениям; содействие ЦНС в регуляции постурального мышечного
тонуса, а также тонуса мышц при движениях сустава; осуществление ин-
гибиторного влияния на поток болевой афферентной активности из сус
тавной рецепторной системы типа IV. Рецепторы типа 1 можно разделить
на статические и динамические механорецепторы.
Г л а в а 6. Нейрофизиология гибкости: невралъная анатомия и физиология
 Суставные рецепторы типа II
Суставные рецепторы типа II
Рецептор типа II представлен крупными коническими частицами, имеющими плотную оболочку. Они располагаются в фиброзной суставной капсуле, однако в более глубоких ее слоях. Каждый кластер обычно состоит из 2—4 частиц. Кроме того, каждый компонент кластера иннервируется ответвлением миел и ниро ванных суставных нервных волокон группы II (диаметром 9-12 мкм). Наибольшая плотность механорецепторов типа II также наблюдается в более дистальных суставах (например, голеностопный), чем в более проксимальных (например, тазобедренный).
С физиологической точки зрения рецепторы типа II, подобно рецепторам типа I, имеют низкий порог. Однако они ведут себя как быстроадапти-руюшиеся механорецепторы и не разряжаются в покое. Следовательно, они полностью пассивны в неподвижных суставах. Рецепторы типа II статически не разряжаются, поскольку их разрядка зависит от скорости. Поэтому их называют динамическими механорецепторами, или механорецеп-торами ускорения. Во время стимулирования каждый кластер осуществляет кратковременный высокочастотный «всплеск» импульсов в течение менее чем 1 с, а нередко и менее 0,5 с. Главная функция этих рецепторов — определение быстрых изменений в движении, таких, как ускорение и замедление.
Суставные рецепторы типа Ш
Механорецепторы типа III представляют собой покрытые тонкой оболочкой частицы, заключенные во внутренние (в пределах суставной капсулы) и внешние (снаружи суставной капсулы) связки большинства суставов. Они отсутствуют в связках позвоночного столба. Это самые большие суставные частицы, которые, подобно нервно-сухожильным веретенам, ведут себя как высокопороговые медленноадаптирующиеся механорецепторы. Они иннервируются миелинированным афферентным аксоном группы I, диаметр которого может достигать 17 мкм.
С физиологической точки зрения, механорецепторы суставных связок имеют высокий порог. Рецепторы типа III являются полностью пассивными в неподвижных суставах и реагируют только при генерировании высокого напряжения в суставных связках.
При стимулировании рецепторы типа III проявляют частоту разрядки, являющуюся непрерывной функцией величины напряжения. Так как это медленноадаптирующиеся рецепторы, разрядка снижается очень медленно (в течение многих секунд), если поддерживается смещение удаленного сустава или тяговое усилие сустава.
Механорецепторы типа Ш имеют две основные функции. Первая — контроль направления движения. Вторая — значительное рефлекторное ингибирование активности некоторых мышц сустава. Таким образом, они могут служить тормозящим механизмом, предупреждающим чрезмерную нагрузку на сустав.
Наука о гибкости
 Суставные рецепторы типа IV
Суставные рецепторы типа IV
В отличие от механорецепторов, нервные окончания типа IV не имеют оболочки. Они делятся на два вида. Окончания типа IVa представлены решетчатыми сплетениями, которые можно обнаружить в жировых прослойках суставов, а также б суставной капсуле. В то же время они полностью отсутствуют в синовиальной ткани, внутрисуставных менисках и суставных хрящах. Рецепторы типа IV6 — это свободные нервные окончания, не связанные со специализированными структурами. Они содержатся во внутренних и внешних связках.
Рецепторы типа IVa и IV6 представляют собой систему болевых рецепторов суставных тканей. В нормальных условиях эти рецепторы являются полностью пассивными. Они проявляют активность, когда суставные ткани, в которых они содержатся, подвергаются существенной механической деформации или химическому раздражению. Примерами химических раздражителей являются такие агенты, как брадикинин, простангландин-Е, молочная кислота, полипептиды, гистамины. Эти вещества появляются в условиях ишемии (нехватка крови) и гипоксии (нехватка кислорода). Как утверждает Уайк (1972):
«Рецепторы категории IV полностью отсутствуют в синовиальной выстилке каждого изученного сустава, а также в менисках коленного и ви-сочно-нижнечелюстного суставов и межпозвонковых дисках. Таким образом, не существует механизма, посредством которого могла бы возникать боль непосредственно а синовиальной ткани или мениске любого сустава, следовательно, хирургическое удаление синовиальной ткани или мениска сустава не означает, таким образом, удаление чувствительной к боли суставной ткани».
Следовательно, подобное хирургическое вмешательство не приведет к снятию боли.
Дата добавления: 2015-05-19; просмотров: 1325;