Нервное волокно как элемент рефлекторной дуги
А. Классификация нервных волокон. Имеется два типа нервных волокон: миелиновые и безмиелиновые. Оболочку безмиелиновых волокон образуют шванновские клетки (леммоциты). Оболочку миелиновых волокон в периферической нервной системе формируют шванновские клетки, а в ЦНС - олигодендроци-ты. Миелиновая оболочка через равные промежутки (0,5-2,0 мм) прерывается, образуя свободные от миелина участки - узловые перехваты Ранвье, протяженность которых в волокнах периферической нервной системы составляет 0,25-1,0 мкм, в волокнах ЦНС их длина достигает 14 мкм. Миелиновая оболочка нервных волокон выполняет изолирующую функцию, обеспечивает более эко--юмное и быстрое проведение возбуждения.
Классификация нервных волокон осуществляется согласно :труктурно-функциональным свойствам. В зависимости от толщи-ш нервных волокон, наличия или отсутствия у них миелиновой оболочки все нервные волокна делят на три основных типа: А, В и (Дж. Эрлангер и X. Гассер). Волокна типа А - это афферентные эфферентные волокна соматической нервной системы, скорость проведения возбуждения 20-120 м/с; волокна типа В - преганг-шонарные волокна вегетативной нервной системы, скорость проведения возбуждения 5-10 м/с; волокна типа С - это в основном постганглионарные волокна вегетативной нервной системы, скорость проведения возбуждения 0,5-2 м/с.
Нервные волокна обеспечивают проведение возбуждения и транспорт веществ, выполняющих трофическую функцию.
Б. Механизм проведения возбуждения по нервному волокну. Биопотенциалы могут быть локальными (местными), распространяющимися с декрементом (затуханием) на расстояние, не
-247 33
превышающее 1-2 мм, и импульсными (ПД), распространяющимися без декремента по всей длине волокна - на несколько десятков сантиметров, например, от мотонейронов спинного мозга по всей длине нервного волокна до мышечных волокон конечностей с учетом самой конечности.
1. Распространение локальных потенциалов. Локальные потенциалы (препотенциал, рецепторный потенциал, возбуждающий постсинаптический потенциал - ВПСП) изменяют мембранный потенциал покоя, как правило, в сторону деполяризации в результате входа №+ в клетку согласно электрохимическому градиенту. В результате этого между участком волокна, в котором возник локальный потенциал, и соседними участками мембраны формируется электрохимический градиент, вызывающий передвижение ионов. В частности, вошедшие в клетку ионы Ыа+ начинают перемещаться в соседние участки, а ионы Ыа+ на наружной поверхности клетки движутся в противоположном направлении. В итоге поляризация мембраны соседнего участка уменьшится. Фактически это означает, что локальный потенциал из первичного очага распространился на соседний участок мембраны. Он затухает на расстоянии 1-2 мм от очага первичной деполяризации, что связано с отсутствием ионных электроуправляемых каналов на данном участке мембраны или неактивацией ионных каналов, продольным сопротивлением цитоплазмы волокна и шунтованием тока во внеклеточную среду через каналы утечки мембраны.
Если возникшая деполяризация мембраны не сопровождается изменением проницаемости потенциалзависимых натриевых, кальциевых и калиевых каналов, такую деполяризацию называют электротонической (ионы перемещаются вдоль клеточной мембраны внутри и вне клетки согласно электрохимическому градиенту). Электротоническое распространение возбуждения - физический механизм, оно характерно для тех фрагментов мембран возбудимых клеток, где нет потенциалзависимых ионных каналов. Такими участками являются, например, большая часть мембраны дендритов нервных клеток, межперехватные промежутки в миели-новых нервных волокнах. Если местный потенциал (рецепторный или ВПСП), распространяясь электротонически, достигает участков мембраны, способных генерировать ПД (аксонный холмик, перехваты Ранвье, часть мембраны дендритов и, возможно, сомы), но его амплитуда при этом не достигнет критического уровня деполяризации, то такой потенциал называют препотенциалом. В его возникновении и распространении частично участвуют потенциалза-висимые ионные каналы, однако при этом нет регенеративной (самоусиливающейся) деполяризации, характерной для ПД. Поэто-
му распространение такого потенциала происходит с затуханием амплитуды. Если локальный потенциал достигает участков мембраны, способных генерировать ПД, и его амплитуда выходит на критический уровень деполяризации, формируется ПД, который распространяется по всей длине нервного волокна.
2. Проведение ПД осуществляется с использованием как физического (электротонического), так и физиологического механизмов. Обязательным условием проведения нервного импульса является наличие на всем протяжении или в ограниченных, но повторяющихся участках волокна потенциалзависимых ионных каналов, ответственных за формирование ПД. В распространении ПД можно выделить два этапа: этап электротонического проведения, обусловленный физическими свойствами нервного волокна, и этап генерации ПД в новом участке на пути его движения, обусловленный реакцией ионных каналов. В зависимости от расположения и концентрации ионных каналов в мембране волокна возможны два типа проведения ПД: непрерывный и сальтаторный (скачкообразный).
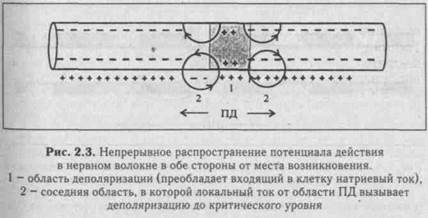
|
Непрерывное распространение ПД осуществляется в безми-елиновых волокнах типа С, имеющих равномерное распределение потенциалзависимых ионных каналов, участвующих в генерации ПД. Возникающий ПД является раздражителем для соседнего участка нервного волокна (рис. 2.3). Таким образом, непрерывное распространение нервного импульса идет через генерацию новых ПД по эстафете, когда каждый участок мембраны выступает сначала как раздражаемый (при поступлении к нему электротонического потенциала), а затем как раздражающий (после формирования в нем нового ПД).
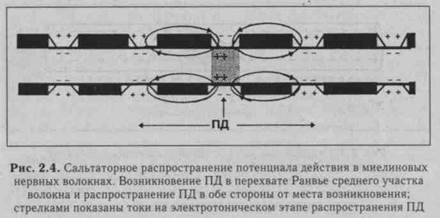
|
Салътаторный тип проведения нервного импульса осуществляется в миелиновых волокнах (типы А и В), для которых характерна концентрация потенциалзависимых ионных каналов только в небольших участках мембраны (в перехватах Ранвье). В области миелиновых муфт, обладающих хорошими изолирующими свойствами, потенциалзависимых каналов почти нет, и мембрана осевого цилиндра там практически невозбудима. В этих условиях ПД, возникший в одном перехвате Ранвье, электротонически (вдоль волокна, без участия ионных каналов) распространяется до соседнего перехвата, деполяризуя там мембрану до критического уровня, что приводит к возникновению нового ПД, т.е. возбуждение проводится скачкообразно (рис. 2.4). Постоянная длина мембраны миелинового волокна достигает 5 мм. Это значит, что ПД, распространяясь электротонически на этом расстоянии, сохраняет 37% своей амплитуды (около 30 мВ) и может деполяризовать мембрану до критического уровня (пороговый потенциал в перехватах Ранвье - около 15 мВ). Поэтому в случае повреждения ближайших на пути следования перехватов Ранвье потенциал действия может электротонически возбудить 2-4-й и даже 5-й перехваты. Саль-таторное проведение ПД, во-первых, более экономично в энергетическом плане, так как возбуждаются только перехваты Ранвье, площадь которых менее 1 % мембраны, и, следовательно, надо меньше энергии для восстановления трансмембранных градиентов Ыа+ и К+, уменьшающихся в процессе формирования ПД. Во-вторых, возбуждение проводится с большей скоростью, чем в безмиелиновых волокнах, так как возникший ПД на протяжении миелиновых муфт распространяется электротонически, что в 107 быстрее, чем скорость физиологического проведения.
В. Характеристика проведения возбуждения по нервным волокнам.
1.Нервные волокна могут проводить возбуждение в двух направлениях. Если в эксперименте нанести раздражение в любом участке нерва, то ПД будет распространяться в обе стороны от участка, на который нанесено раздражение.
2. Возбуждение проводится изолированно в каждом нервном волокне. Это обусловлено тем, что петли тока в межклеточной жидкости ствола, имеющей низкое сопротивление, почти не проникают в невозбужденные нервные волокна из-за большого сопротивления их оболочек. Изолированное проведение импульсов по нервным волокнам обеспечивает точное афферентное и эфферентное влияния функционально разнородных волокон нерва. Однако при одновременном раздражении большого числа нервных волокон возможно возбуждение других - прилежащих волокон и усиление нервных влияний.
3. Большая скорость проведения возбуждения, достигающая 120 м/с. Передача возбуждения по нервным волокнам является наиболее скоростным из известных способов передачи информации на значительные расстояния в организме. Скорость передачи гуморальных влияний ограничена скоростью кровотока (полный кругооборот частицы крови совершают за 22 с).
4. Малая утомляемость нервного волокна. При нормальной доставке кислорода и питательных веществ проводящий возбуждение нерв практически неутомляем. Это обусловлено тем, что при проведении одного ПД по нервному волокну используется всего лишь одна миллионная часть запасов трансмембранных ионных градиентов и, следовательно, нужны небольшие количества АТФ для восстановления ионных градиентов. Расход энергии в нервном волокне на единицу массы примерно в 16 раз меньше, чем в целом организме в условиях основного обмена, и в миллион раз меньше, чем в работающей мышце.
5. Возможность функционального блока:проведения возбуждения при морфологической целостности волокон (Н. Е. Введенский). Нарушение физиологической непрерывности нервных волокон возникает при действии электрического тока, анестетиков, воспалении, гипоксии, охлаждении. После прекращения действия этих факторов проведение возбуждения по волокнам нерва восстанавливается. Причиной блока проведения возбуждения является инактивация Ыа-каналов.
Г. Аксонный транспорт. Основная масса веществ (ферментов, структурных белков, полисахаридов, липидов) образуется в теле нейрона, а используются они в различных его отделах. Для
транспорта веществ путем диффузии на расстояние, равное максимальной длине аксона (около 1 м), потребовалось бы 50 лет! Транспорт в отростках нейрона лучше изучен в аксонах и получил название аксонного транспорта, с помощью которого осуществляется трофическое влияние в различных участках нейрона и на иннерви-руемые клетки. Транспорт веществ в дендритах осуществляется из тела клетки. Имеется быстрый - перемещение различных веществ в двух направлениях (200-400 мм/сутки) - и медленный аксон-ный транспорт - передвижение всего столба аксоплазмы в антег-радном направлении со скоростью 1-2 мм/сутки, что соответствует скорости роста аксона в онтогенезе и при регенерации.
Аксонный транспорт играет важную роль также и при регенерации поврежденных нервных волокон.
Дата добавления: 2015-05-16; просмотров: 1082;