Детерминанты половых клеток
Будущие половые клетки – гоноциты иногда можно отличить уже на очень ранних стадиях развития. Классическим примером является аскарида, у которой после первого деления яйца в одном из двух бластомеров теряются концевые участки хромосом и способность стать клеткой зародышевого пути сохраняет только второй бластомер. Аналогичным образом происходит потеря нескольких целых хромосом (или даже наборов хромосом) у некоторых насекомых.
У большинства животных раннее обособление гоноцитов связано не с потерей части ДНК, она целиком сохраняется во всех клетках зародыша. Но в цитоплазме многих яиц обнаружены особые частицы – половые детерминанты. Природа этих детерминантов, их структура и механизм действия известны плохо. Вероятно, у разных видов они различны. В некоторых случаях это частицы, состоящие из РНК и белка. У рачка циклопа половые детерминанты попадают в один из двух первых бластомеров, при втором делении оказываются в одной из четырех клеток и т. д. После шестого деления, когда зародыш уже состоит из многих десятков клеток, детерминанты оказываются всего в двух клетках – гоноцитах.
У дрозофилы первые восемь делений ядер происходят в центре яйца без деления цитоплазмы. Но уже после первых делений ядер два из них сдвигаются к заднему концу яйца, где находятся половые детерминанты, – эти два ядра дадут начало половым клеткам. Это можно показать экспериментально. Если задний конец яйца перевязать или инактивировать половые детерминанты ультрафиолетом, муха не будет иметь половых клеток. Если же в эту область яйца инъецировать ядра из другого зародыша, муха будет иметь половые клетки той генетической линии, откуда были взяты ядра.
В яйцах лягушки половые детерминанты находятся на вегетативном полюсе. Если эту часть яйца облучить ультрафиолетом, то из него разовьются внешне нормальные животные, в половых железах которых, однако, не будет половых клеток.
Обособление гоноцитов у птиц и млекопитающих обнаруживается существенно позже. Гоноциты теплокровных находятся вне собственно зародыша, в так называемых внезародышевых частях. Потом гоноциты у зародышей птиц переносятся в зачатки половых желез по кровеносным сосудам, а у млекопитающих вползают туда посредством собственных амебоидных движений. Их движение в железу стимулируется особым веществом.
У гидры и других кишечнополостных и у некоторых червей половые клетки не проходят стадию гоноцитов, а в течение всей жизни образуются из специальных малодифференцированиых і‑клеток, которые служат общим резервом для различных дифференцировок.
В чем биологический смысл особого происхождения гоноцитов? Самое разумное объяснение заключается в том, что такая их особая судьба позволяет лучше защитить клетки зародышевого пути от «нежелательных» влияний, которые могли бы изменить их ДНК. А это, в свою очередь, позволяет сделать очень важное предположение о том, что во время развития органов и тканей всегда или достаточно часто, закономерно или случайно происходят какие‑то необратимые изменения ДНК. Ниже мы еще вернемся к этому принципиальному вопросу. У растений и у низших многоклеточных, у которых образование половых и соматических клеток не обособлено друг от друга, вероятно, нет и подобных изменений ДНК.
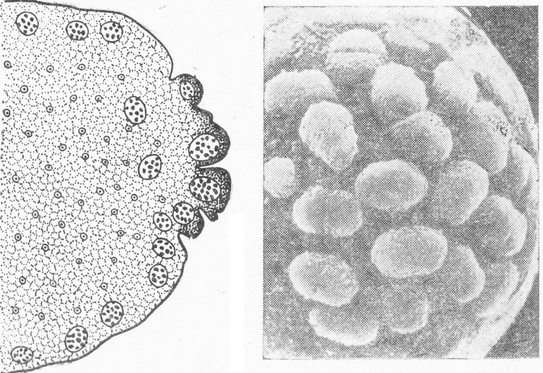
Особое происхождение половых клеток – полярные клетки в раннем развитии насекомых
Слева – рисунок среза задней части яйца. Видны ядра, расположенные в поверхностном слое цитоплазмы, и на самом заднем полюсе – более крупные ядра, окруженные темной цитоплазмой, только из них образуются генеративные клетки – яйца и сперматозоиды; справа – электронная фотография заднего полюса яйца. Полярные клетки как бы лежат на поверхности. Некоторые из них делятся
Дата добавления: 2015-05-08; просмотров: 1060;