III. Суггестия
В главах 4 и 5 мы рассмотрели физиологические явления тормозной доминанты и имитативного рефлекса и показали, что их встреча в высшей нервной деятельности семейства троглодитид должна была породить высоко развитое явление интердикции. Наибольшую ступень интердикции мы назвали генерализованной интердикцией: имитационное провоцирование некоторого одного действия парализует возможность каких бы то ни было других действий (очевидно, за исключением функционирования автономной нервной системы) и это состояние парализованности, вероятно, может продолжаться долго даже после прекращения действия данного имитатогенного агента.
Тем самым высшую форму интердикции можно было бы считать низшей формой суггестии: это уже не торможение лишь того или иного отдельного действия, но навязывание некоего состояния, допустим, типа каталепсии. Однако таков лишь зачаток суггестии, ибо под ней (под "внушением") понимается возможность навязывать многообразные и в пределе даже любые действия. Последнее предполагает возможность их обозначать. Между этими предельными рубежами умещается (развивается) явление суггестии. Нас прежде всего интересуют его первые шаги.
Надо напомнить еще раз коренной тезис, что вначале, в истоке, вторая сигнальная система находилась к первой сигнальной системе в полном функциональном и биологическом антагонизме. Перед нашим умственным взором отнюдь не "добрые дикари", которые добровольно подавляют в себе вожделения и потребности для блага другого: они обращаются друг к другу средствами инфлюации, к каковым принадлежит и суггестия, для того чтобы подавлять у другого биологически полезную тому информацию, идущую по первой сигнальной системе, и заменять ее побуждениями, полезными себе. Это явление инфлюации, в том числе суггестии, не имеет никакого отношения к гносеологии. Сколько обвинений в идеализме и мистицизме было обрушено на вывод Леви-Брюля о "прелогическом мышлении", тогда как это явление действительно существовало на заре истории и проявлялось впоследствии, но только оно не было "мышлением": оно было подавлением первосигнального (еще единственно верного тогда) способа отражения окружающей среды и системой принудительного воздействия на поведение друг друга. Да, оно принадлежит не к гносеологии, а к онтологии. Это только взаимодействие особей. Тут нет отношения субъекта к объекту, а есть лишь отношение организма к организму. "Прелогическая стадия" ничуть не угрожает логике: тут логике еще решительно нечего было делать. Мы убедимся, что и антилогика в самом деле гораздо раньше объявилась в этой сфере отношений между индивидами, чем логика в сфере человеческого познания. Но о генезисе логики мы будем говорить только в конце этой главы (и, тем самым, этой книги).
Развитие второй сигнальной системы у людей ни в коем случае не было следствием разрастания общего объема головного мозга по сравнению с объемом (весом) тела и прямо никак не связано с этим процессом энцефализации (церебрализации) в филогении троглодитид. Во-первых, в биологической эволюции вообще налицо тенденция увеличения мозга (независимо от размеров тела), и по этому показателю прямоходящие высшие приматы, включая человека, вовсе не оказываются специфичными: степень энцефализации увеличивается у всех ископаемых млекопитающих. По степени развития неокортекса ("новой коры"), т. е. по степени "неоэнцефализации", можно выделить весь отряд приматов из общей родословной млекопитающих, но невозможно отдифференцировать собственно человеческую линию 36. Во-вторых, разрастание объема головного мозга в эволюции семейства троглодитид было прямым морфологическим следствием прямохождения и прямым биохимическим следствием плотоядения, т. е. повышенного усвоения протеина 37; эти два фактора влекли за собой широкий размах индивидуальных вариаций объема мозга, иначе говоря, делали структурно и гистологически возможными колебания в сторону повышенной массы мозга, а естественный отбор закреплял эти отклонения, несомненно, потому, что они были биологически выгодны. По Боне, это разрастание мозга происходило в том же темпе, как и другие морфологические трансформации млекопитающих в плейстоцене. Но у Homo sapiens средний размер и вес головного мозга не возрастал и не возрастает сравнительно с поздними палеоантропами. При этом размах индивидуальных вариаций данного признака у Homo sapiens весьма увеличился сравнительно с палеоантропами, нередко встречается объем мозга, значительно превышающий среднюю величину, но ничто не закрепляет этих отклонений: они биологически нейтральны и средняя величина остается неподвижной для черепов любого времени с верхнего палеолита до наших дней. Между тем речевая функция мозга в корне отличает неоантропа от палеоантропа.
Как видно, социальность и разум человека никак прямо не коррелированы с тотальной величиной его головного мозга.
Зато корреляция начинает проступать, когда измерению подвергаются по отдельности длиннотный, широтный и высотный диаметры роста головного мозга в филогенетической цепи: шимпанзе австралопитек археоантроп палеоантроп неоантроп. До неоантропов наиболее интенсивно увеличивался тотальный длиннотный размер (за счет роста задних областей); напротив, только мозг неоантропов дает интенсивный сдвиг высотного диаметра: в лобной и теменной долях эпицентры роста перемещаются из нижних отделов в верхние, происходит, как мы уже отмечали, усиленный рост префронтальной области, а также поднятие и выравнивание поверхности мозга в его своде 38.
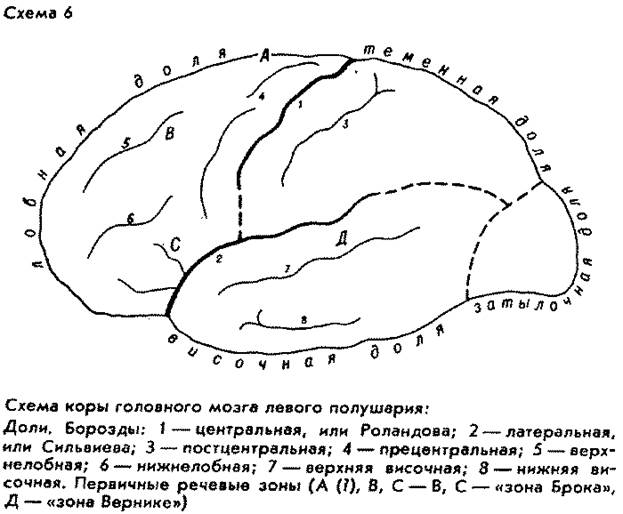
Интересно, что как раз в самой высотной, и тем самым весьма молодой, точке свода нашего мозга предположительно локализуется самый исходный, "инициальный" очажок речевой функции человека (см. схему коры мозга). Правда, вопрос пока является дискуссионным и открытым. Это так называемая "речевая зона Пенфильда", или "верхняя речевая кора", якобы зона начальной речевой активизации 39. Эту зону Пенфильд и Роберте якобы обнаружили при оперативных исследованиях эпилепсии в так называемом дополнительном моторном поле на медиальной (внутренней, т. е. обращенной к другому полушарию) поверхности в задних отделах верхней лобной извилины. Некоторые авторы считают это прочно установленным фактом 40, другие при применении иных нейрохирургических методов не обнаруживают нарушений речевых функций при поражениях этой зоны (А. Р. Лурия) или замечают воздействие всего лишь на оттенки интонации и модуляции. Впрочем, последнее не служило бы решающим возражением: кто знает, может быть, так и рождалась первичная дифференциация звуков-знаков. Не будем пока ни принимать, ни отбрасывать гипотезу Пенфильда Робертса. Заметим лишь, что, если бы указанная зона в самом деле все-таки оказалась начальным очажком второй сигнальной системы, было бы поразительно, что он расположен в самой высотной части, т. е. на самом молодом верхнем крае коры мозга Homo sapiens. В пользу второсигнальной принадлежности этого образования (если принять описание его Пенфильдом и Робертсом) говорит его односторонняя локализованность в доминантном полушарии (в левом у правшей), тогда как близко расположенные центры локализации находятся в обоих полушариях.
Интересно, что довольно высоко в структуре коры головного мозга Homo sapiens, впереди от средней части прецентральной извилины, лежит и участок, управляющий элементарными графическими действиями. Это свидетельствует, что они возникают на весьма ранних стадиях генезиса второй сигнальной системы. Люди начальной поры верхнего палеолита уже "рисовали", еще не вполне умея "разговаривать", если применять эти термины в современном смысле. Туда, в эту древность, уходят корни письменной речи. Зато когда иные археологи фантазируют, будто они обнаружили "рисунки" мустьерцев (палеоантропов, неандертальцев), мы уверенно можем считать это исключенным в той же мере, как заявку на изобретение перпетуум-мобиле: в мозге палеоантропов еще не было той высотной части, где находится центр, управляющий графическими действиями; они не могли носить и неуправляемого двигательно-подражательного характера (какие осуществляют шимпанзе), так как подражать в "рисовании" еще было тогда некому.
Еще далее вперед от самой высотной части свода головного мозга Homo sapiens (где сходятся лобная и теменная доли) лежит тот префронтальный отдел, о роли которого, в особенности его верхней формации, мы уже говорили в связи с постановкой проблемы происхождения второй сигнальной системы. Это образование присуще только Homo sapiens.
Можно сказать, что это главное морфологическое звено второй сигнальной системы. Отсюда по исключительно богатым нервным путям и контактам происходит возбуждение, активация (с помощью сетевидной, или ретикулярной, формации) отвечающих "задаче" или "намерению" двигательных центров мозга и одновременное торможение всех других двигательных центров, не идущих к делу 41.
Эволюционная морфология мозга не может ничего прямо сказать о развитии функциональной асимметрии полушарий (доминантность субдоминантность, правшество левшество). Вообще говоря, асимметрия как один из аспектов парности работы полушарий головного мозга обнаружена у животных: симметричные центры двух полушарий могут в данный момент находиться во взаимном антагонизме, т. е. в одном полушарии выполнять функцию возбуждения, во втором торможения 42. Но у человека асимметрия закреплена: лишь кора одного полушария, обычно левого (у правшей), управляет всей второсигнальной функцией. Афазиологией доказана левосторонность управления динамикой как речевой, так и рече-мыслительной деятельности в лобных долях. Однако пока недостаточно подчеркивается, что вся эта односторонность управления второй сигнальной системой, т. е. локализация всех прямо причастных к ней зон и центров в "доминантном" полушарии, означает сопряженное торможение центров, управляющих неречевыми движениями преимущественно с противоположного, "субдоминантного" полушария, но отчасти с того же 43. Если так, можно существенно переакцентировать обычное восприятие понятий "правшество", "левшество": дело не в том, что у "правши" правая рука (и другие органы) обладает некими повышенными возможностями, а в том, что у него, напротив, приторможена, снижена левосторонняя моторика (в первую очередь руки), у "левши" наоборот. Есть немало оснований ограничить недавнее слишком абсолютное представление о двигательном "правшестве" и "левшестве": в большинстве случаев то и другое как-то смешано в индивиде. Но по некоторым данным онтогенеза можно предполагать, что в филогенезе асимметрия прошла три фазы: правшество левшество снова правшество; поэтому их следы у нас могут наслаиваться друг на друга. Что же мы обнаружили, выясняя древнейшие уровни, "исходные рубежи" второй сигнальной системы с помощью эволюционной морфологии мозга, афазиологии, генетической психологии? Мы обнаружили, что древнейшие зоны речевой деятельности возникают в моторной (двигательной), а не сенсорной (чувствующей) области коры. Это и отвечает выдвинутому выше тезису, что вторая сигнальная система родилась как система принуждения между индивидами: чего не делать; что делать. Мы получим дальнейшее подкрепление этого тезиса, перейдя от тотальных параметров, характеризующих особенности головного мозга Homo sapiens, к эволюционному сравнению состояния отдельных долей, извилин и борозд, поскольку они отразились на эндокранах ископаемых предковых форм, и к частным видам афазий.
Подчас антропологи принимают намеченное на эндокране питекантропа образование на месте так называемой "зоны Брока" в нижней лобной извилине за свидетельство присутствия у него речи, исходя из того, что у современного человека повреждения этой зоны вызывают нарушение двигательной (произносительной) речевой функции моторную афазию. Со своей стороны повреждения "зоны Вернике" в первой височной извилине вызывают сенсорную афазию нарушение восприятия чужой слышимой речи. Но речь не просто наличие двух устройств передающего сигналы и принимающего их, тем менее одного из двух; недостаточно и двусторонней связи между ними, т. е. единства слухо-двигательного анализатора. Допустим, питекантропы могли издавать и принимать разные крики; это не членораздельность, не речевые знаки в смысле современной науки. В. В. Бунак на основе анатомии периферических органов речи гортани, челюстного скелета установил, что у форм, предшествовавших Homo sapiens, не могло быть членораздельной синтагмической речи 44 (дедукция какой-то якобы предшествовавшей, "нечленораздельной речи" не удалась она противоречит всей аксиоматике науки о речи); в 1971 г. появилось сообщение, что Ф. Либерман (ун-т Коннектикут) изготовил искусственный макет гортани неандертальца (шапелльца) и получил экспериментальное физическое подтверждение того, что она не могла бы производить членораздельных звуков 45.
Однако суть дела вовсе не в этой физико-акустической стороне проблемы. Она попала в центр внимания только тех авторов, которые не в курсе современной лингвистики, как и афазиологии. Поражение "зоны Вернике" в височной доле вызывает не потерю слуха или возможности различать звуки, нарушает не акустическую основу восприятия речи, словом, порождает не "фонетическую глухоту", но "фонологическую глухоту" невозможность правильно узнавать фонемы как смыслоразличающие элементы речи. Наука фонология, отделившаяся от фонетики (за которой осталась физиология звукообразования и звуковосприятия), изучает наличные во всяком языке звуковые "букеты", которые психика человека расценивает как один и тот же элемент, сколь бы ни отличались друг от друга эти звуки в плане акустическом, и которые имеют и сохраняют свою определенность только по противопоставлению другому такого же рода "букету" в той же артикуляционно-фонетической группе 46. Именно в этом проявляется тут "чрезвычайная прибавка" человека в возможности отождествлять звуки, высоко различные акустически (ср., например, весьма искаженное "повторение" маленьким ребенком слов взрослого), как и, напротив, в возможности делать эти звуки принципиально нетождественными даже при их немалой акустической близости посредством оппозиции, по природе столь же абсолютной, как и нейрофизиологические явления возбуждения и торможения.
Что касается не восприятия речи, а говорения, то оказалось, что и двигательная (моторная) сторона речевой деятельности раздвоена точно так же. Поражения управляющих ею зон в коре мозга, в том числе нижней лобной извилины, нижнего отдела прецентральной извилины, нижнего отдела теменной доли, приводят либо к эфферентной моторной афазии нарушению функции противоположной фонематической, а именно функции не разделения фонем, а их связности, плавности, слияния хотя бы и различных звуковых единиц в одну единицу, например в слог; либо к афферентной моторной афазии смешению, неразличению фонем, в частности, близких по артикуляции. В последнем случае перед нами патология не самой артикуляции звуков органами речи, а психо-физиологического механизма контроля этих движений по фонологическим меркам, иначе говоря, психо-физиологического механизма обратной коррекции речевых движений 47.
Выходит, что "сенсорная афазия" и "моторная афазия" представляют собою два свидетельства одного и того же факта появления в мозге Homo sapiens на определенном этапе его формирования принципиально нового уровня реагирования. Важно, что оба они и в анатомо-физиологическом смысле тесно соединены. Нижние отделы лобной и теменной долей (очаги моторных афазий) близко примыкают к переднему отделу височной доли (очагу сенсорной афазии). Только будучи по существу единым аппаратом, они могут осуществлять эхолалическую (речеподражательную) операцию, лежащую глубоко в основе всей нашей речевой, а тем самым и речемыслительной деятельности: непроизвольное повторение слышимого, причем не на акустико-фонетическом (не как у попугаев или скворцов), а именно на фонологическом уровне. В норме эта операция у нас редуцирована, так что зарегистрировать ее могут только тончайшие электрофизиологические приборы 48, но при поражениях или функциональных расстройствах в коре, т. е. когда подавляющие ее позднейшие нервные образования вышли из строя, она выступает с полной наглядностью и назойливостью. Врач говорит больному "встаньте", тот повторяет "встаньте", но не встает. Эхолалическая реакция на речь не несет никакой смысловой нагрузки. Показано, что это явление характерно для ранней стадии освоения речи ребенком младшего возраста, а также для различных нервных расстройств (неврозов), в том числе истерии 49. Тот факт, что этот фундаментальный механизм речевой деятельности протекает у нас необычайно быстро, следовательно, по простейшим нейронным путям, свидетельствует о его особой древности, в некотором смысле даже первичности в эволюционном становлении второй сигнальной системы у неоантропов.
Этому соответствует указанная анатомическая связь органов, или зон фонологического анализа и контроля, в коре, в частности близость и взаимосвязь нижнелобной и височной долей. Если у австралопитеков лобная и височная доли плотно примыкали друг к другу, то у синантропов и палеоантропов они были резко разделены довольно широкой, глубокой и узкой сильвиевой ямкой, а у неоантропов края ее снова соединились, однако переместившись вперед и при новых очертаниях всей височной доли 50. Современные знания о работе, выполняемой у нас U-образным изгибом коры в глубине височной ямки, недостаточны для интерпретации этого крутого морфологического преобразования. Однако можно с известной долей уверенности предполагать, что указанный изгиб осуществляет прямую и кратчайшую нейронную связь между "зоной Брока" и "зоной Вернике" и тем самым обеспечивает эхолалическую (речеподражательную) подоснову второй сигнальной системы.
Прежде чем в нашем обзоре коры двинуться дальше по направлению от лба к затылку, поищем ответ на вопрос: какая из только что рассмотренных двух взаимодействующих речевых зон (лежащих примерно по названному направлению) эволюционно старше? Афазиология располагает наблюдением, подсказывающим ответ. У больных с моторной афазией более или менее нарушена глагольная сторона экспрессивной речи, тогда как при сенсорной афазии страдают имена существительные, отчасти прилагательные 51. Мы уже знаем локализацию этих афазий. По-видимому, тем самым глагольная фаза второй сигнальной системы ("нижнелобная" и "нижнетеменная") оказывается старше, чем предметно-отнесенная ("височная"). И в самом деле, многие лингвисты предполагали, что глаголы древнее и первичнее, чем существительные. Эту глагольную фазу можно представить себе как всего лишь неодолимо запрещающую действие или неодолимо побуждающую к действию. В таком случае древнейшей функцией глагола должна считаться повелительная. Можно ли проверить эту гипотезу? Да, несколько неожиданным образом: демонстрацией, что повелительная функция может быть осуществлена не только повелительным наклонением (например, начинайте!), но и инфинитивом (начинать!), и разными временами прошедшим (начали!), настоящим (начинаем!) и будущим (начнем!), даже отглагольным существительным (начало!). Словно бы все глагольные формы позже разветвились из этого общего функционального корня. И даже в конкретных ситуациях множество существительных употребляется в смысле требования какого-либо действия или его запрещения: "огонь!" (стрелять!), "свет!" (зажечь), "занавес!" (опустить), "руки!" (убрать, отстранить). Последний пример невольно заставляет вспомнить, что Н. Я. Марр обнаружил слово "рука" в глубочайших истоках больших семантических пучков чуть ли не всех языков мира: "рука" означала, конечно, не предмет, а действие. Вот совсем другая подкрепляющая гипотезу демонстрация. Знаменитый путешественник В. К. Арсеньев записывает коверканную русскую речь проводника гольда Дерсу Узала, где почти все глаголы, а то и существительные (еда "кушай") употреблены в повелительном наклонении: рассматривая следы "Давно одни люди ходи. Люди ходи кончай, дождь ходи"; оставляя на стоянке запас "Какой другой люди ходи, балаган найди, сухие дрова найди, спички найди, кушай найди пропади нету" 52. Несомненно, это воспроизводит некоторые архаизмы самого гольдского языка. Мы встречаем этот же курьез и в ломаной русской речи лиц некоторых других национальностей (например, "не понимай"). Итак, допуская, что древнейшими словами были глаголы, мы вместе с тем подразумеваем, что глаголы-то были лишь интердиктивными и императивными, побудительными, повелительными.
Теперь завершим наше путешествие по центрам и зонам второй сигнальной системы коры головного мозга. На ближних к рассмотренным участках коры расположены и центры, которые при поражении верхних слоев клеток управляют парафазиями, т. е. непроизвольными деформациями воспроизведения звуков и слов их перестановками, подменами по противоположности звуков или по ассоциации слов. Это явление очень важно, оно может считаться самоотрицанием (или просто отрицанием) эхолалии антиэхолалией.
Дальше к задним частям мозга, на стыках височной области с теменной и затылочной, как и на стыке двух последних, находятся весьма миниатюрные (с орешек), но и весьма важные очажки второсигнального управления осведомлением о внешней среде и действиями в ней (гнозисом и праксисом). Некоторые авторы соответственно усматривают именно в этих тонких корковых образованиях собственно "речевые органы" центральной нервной системы. Наконец, на стыках этих трех областей задней надобласти коры исследователи цитоархитектоники обнаружили у человека в отличие от животных особенно сложные многоклеточные ядра. Допустимо предположить, что все это сформировалось как морфофункциональные механизмы эффективного исполнения суггестии, в частности императивной (предписывающей выполнить те или иные действия в среде).
Итак, мы обогнули все доминантное полушарие человека. Мы начали с предположительной, или, если угодно, сомнительной, "верхней речевой коры", которой приписывается инициальная роль в речевом акте, твердо смогли опереться на современные обильные научные данные о функциях верхних передних отделов лобной доли, благодаря чему интердикция заняла надлежащее ей место исходной ступени в генетическом рассмотрении второй сигнальной системы, прошли через классические сенсорные и моторные формации и закончили в задней надобласти коры. В литературе, касающейся антропогенеза, встречается лишь обратная схема. Так, выдающийся невропатолог-эволюционист Е. К. Сепп усматривал исходный пункт развития специфически человеческих функций и структур мозга в координации анализаторов задней надобласти коры, якобы независимо от второй сигнальной системы, от общения людей обеспечивавшей тонкие трудовые действия каждой единичной человеческой особи 53. Очень жаль, что В. И. Кочеткова поддалась влиянию этого построения, ориентированного на эволюцию отдельно каждого индивида, когда ее собственные научные достижения давали основание для его опровержения. Увы, почти всякий современный автор, рассуждая о роли "труда" в становлении человека, подразумевает именно единичную особь, манипулирующую с материальными предметами 54, т. е. начинает с задней части мозга.
Мы произвели все это путешествие по речевым зонам мозга одновременно и как путешествие по истории становления второй сигнальной системы неоантропа в период его дивергенции с палеоантропом. Каждый наш шаг по небольшой поверхности есть шаг и в длительном времени. Это ступени развития феномена суггестии. Оно в целом укладывается между двумя рубежами: возникает суггестия на некотором предельно высоком уровне интердикции; завершается ее развитие на уровне возникновения контрсуггестии. Но какие сложные трансформации на этом пути! С другой стороны, как сложны на этом пути и взаимодействия между преобразованиями нервных функций и преобразованиями мозговых формации, тканей и клеток, так же как периферийных органов речи! В соответствии с твердо установленным биологами законом и здесь функция и морфология менялись вместе, во взаимодействии. К примеру, возникновение фонологической дифференциации и группировки звуков, вероятно, дало огромный толчок обогащению, прогрессу нервных центров, ведающих и простой акустико-фонетической артикуляцией, так же как скелетно-мышечных органов произношения звуков. Но и обратно, некоторые вариации этих тканей или органов благоприятствовали зарождению фонологической функции. Мозг перестраивался вместе, в единстве с генезисом второй сигнальной системы. В том числе, как мы знаем, он расплатился утратой немалой части затылочной (в основном зрительной) доли; можно предположить, что мы тем самым лишились принадлежавшей неандертальцам способности хорошо и видеть, и передвигаться в полутьме, замечать малейшие помехи и опоры для локомоции и т. д.
На пути развития существенно менялась сама природа суггестии. Приметим, что пока мы оперировали суммарными, или тотальными, макропараметрами человеческого мозга, механизмы влияния ( инфлюации) еще можно было трактовать в рамках понятия интердикции. Напротив, когда в конце обзора мы подошли к детальным микроструктурам на стыках долей, в том числе долей задней надобласти височной, теменной и затылочной, дело пошло о таких механизмах речевого воздействия, которые требуют понятия императива (предписания, прескрипции). Таким образом и в этом отношении обнаруживается существенное и глубокое различие, даже противоположность двух крайностей, между которыми совершалось эволюционное поступательное движение, или, что то же, становление феномена суггестии.
Не будем скрывать от себя, что самое трудное объяснить первый шаг и, напротив, чем ближе к завершению данного процесса, тем более очевидный или вероятный может быть предложен анализ. В самом деле, ведь вот же феномен интердикции, даже в ее развитой, или генерализованной, форме, мы оставили в царстве первой сигнальной системы допустили, что для него не требуется того высокого лба, который отличает Homo sapiens. А в то же время допускаем, что именно с функции интердикции начинается восходящий ряд феномена суггестии, и локализуем эту функцию как раз в специфических для второсигнального уровня и для мозга Homo sapiens верхних лобных формациях. Логика требует считать, что сама интердикция претерпела при этом качественное изменение: только в таком случае соблюдается принцип биологической непрерывности, хотя бы посредством инверсии. Иными словами, мы лишь свели до минимума тот участок перехода от первой сигнальной системы ко второй (тем самым от животного к человеку), который, вероятно, поколения специалистов будут исследовать.
Ниже предлагается все же рабочая модель этого метаморфоза интердикции, превращения ее из одного качества в другое, противоположное. Интердикция I: генерализованный тормоз, т. е. некий единственный сигнал (не обязательно думать, что он звуковой: вероятнее, что это движение руки), тормозящий у другой особи, вернее, у других особей, любое иное поведение, кроме имитации этого сигнала. Интердикция II: некий сигнал, специально тормозящий этот генерализованный тормоз ("интердикцию I"), вызывая имитацию на себя, т. е. провоцируя ту деятельность, которая служит тормозной доминантой для действия "интердикция I". Однако это не может мыслиться просто как движение по кругу, как повторение начальной схемы. Это спираль, выход на новый уровень. Так, правдоподобно, что этот новый сигнал сам был полиморфным: звуковым, но не каким-либо отдифференцированным звуком, а любыми издаваемыми звуками, т. е. адресованным звуком вообще. В таком случае его адресованность состояла в том, что он был действенным, только если кто-то осуществлял "интердикцию I".
Иначе говоря, мы допускаем гипотезу, что "интердикция II" представляла собою звукоиспускание более или менее генерализованное по физиологической природе и диффузное с точки зрения лингвистической. Конечно, выражение "любой звук" на деле, вероятно, требовало бы ограничения, так как возможно, что те или иные звуки, очень специализированные по механизму испускания (скажем, свист), могли оставаться вполне сепаратными в отношении данного комплекса. С другой стороны, мы можем предположить, что троглодитиды относятся к числу тех нечеловеческих приматов, которые располагали бедным набором звуков; как известно, среди обезьян есть и очень богатые различными звуками (в том числе особенно среди низших), и очень бедные. Мы навряд ли ошибемся, сказав, что ближайшие предки людей принадлежали к числу последних, и даже в крайней степени.
Это диффузное звукоиспускание, вызывая неодолимым (роковым) образом имитацию, парировало "интердикцию I". Оно не имело никакого иного биологического назначения. Оно лишь освобождало какое-либо действие от примитивного "нельзя" снимало запрещение. Можно сказать: оно запрещало запрещать что и было самым первым проблеском гоминизации животного.
Следующим шагом не могло быть ничто иное, кроме отрицания и этого отрицания. Диффузный комплекс звуков теперь делится на два, составляющих оппозицию друг другу по характеру артикуляции, или звукоиспускания. Каждый из двух остается внутри в высокой мере диффузным. Однако дифференциация между ними настолько определенна, что один способен служить тормозной доминантой и сигналом интердикции, т. е. неким физиологическим "наоборот" в отношении другого. Мы не можем пока знать, обеспечена ли эта бинарная оппозиция физиологической противоположностью звуков при вдыхании (инспирации) и выдыхании (экспирации) или достигнутой несовместимостью некоторых приемов артикуляции согласных. Так или иначе, в этом раздвоении "противная сторона" обрела средство парализовать, затормаживать то самое ("интердикцию II"), чем на предыдущем этапе парализовали ее собственное тормозящее устройство "интердикцию I". Если угодно, пусть назовут это средство "интердикция III", однако такой новый термин был бы излишен, так как мы лишь раскрыли теперь генетическое содержание понятия "суггестия".
Впрочем, в поисках поясняющих слов можно было бы обозначить, хотя и неточно, три описанных ступени тремя терминами из современного языка, следовательно, несущими сейчас существенно иной смысл: I "нельзя", II "можно", III "должно". Последнее и есть прескрипция. В этом случае партнеру ("противнику") предписывается, или навязывается, не нечто внутренне неопределенное, каково "звукоиспускание вообще", но нечто имеющее определенность внутри данного качества. Можно расположить эти три явления филогенетически, как относящиеся к истории дивергенции неоантропов с палеоантропами. I. "Интердикция I" есть высший предел нервных взаимодействий между особями еще в мире палеоантропов. II. "Интердикция интердикции" ("интердикция II"), т. е. самооборона, есть характерное нервное взаимодействие в механизме самой дивергенции: взаимодействие между Homo sapiens и Troglodytes. III. "Интердикция интердикции интердикции" есть перенесение отношений, характерных для дивергенции, в мир самих неоантропов в плоскость взаимодействий между особями и группами Homo sapiens. В этом последнем случае потенциал дальнейших осложнений безграничен.
По-видимому, надлежит думать, что два оппозиционных звуковых комплекса вполне реципрокны: каждый может служить тормозной доминантой в отношении другого. Но нет причин думать, что в той или обратной роли один комплекс нейрофизиологически был закреплен лишь за одними особями, второй за другими. С чисто биологической точки зрения все неоантропы могли бы пользоваться попеременно, т. е. в равной мере, обоими противоположными звуковыми комплексами для обеих противоположных функций. Однако, возможно, это не было так: мы находимся где-то у истоков бинарной, или дуальной, группировки людей, т. е. за частью их закрепляется в активной функции один комплекс, за частью второй. Это можно уподобить математическим положительному и отрицательному знакам или противоположным электрическим зарядам. Сейчас нам важно лишь то, что у этого закрепления нет ни малейшей биологической, в частности нейрофизиологической, детерминированности: детерминирован лишь сам факт оппозиции, восходящий к полярности возбуждения и торможения. Но в предлагаемой схеме мы уже имеем дело с начатком "языковых", или "культурных", оппозиции, каковые могут быть в дальнейшем сколь угодно обширными и множественными без всякого дальнейшего развития этой нейрофизиологической основы. Следует только помнить, что они никогда не могут существовать без нее.
Кратко описанная исходная бинарность, или дуальность, является лишь мысленной реконструкцией с помощью нейрофизиологии (с помощью теории тормозной доминанты), но она не поддается проверке прямым наблюдением ни над детьми, ни над археологическими древностями, ни над этнологическими или лингвистическими "следами". Н. Я. Марр, создав небывалый инструментарий палеолингвистики, хотя и пробурил историческую толщу, все же смог полуинтуитивно нащупать лишь последующий пласт: членение не на два, а на четыре звуковых комплекса, внутренне диффузных, что, может быть, отвечает бинарности, пересеченной новой бинарностью.
Однако именно поэтому на данном уровне мое изложение может быть прервано. Я призываю читателя обратиться заново к могучим, хоть и недостаточно строгим, прозрениям Марра. Теперь, когда его выводы о четырех древнейших лингвистических элементах, как и другие палеолингвистические находки, оказались в известном соответствии с результатами, достигнутыми совершенно иным, биологическим, методом, они снова обрели право на внимание. Эта физиологическая опора и проверка едва не пришла еще при его жизни: исследовательская мысль И. П. Павлова и всей "могучей кучки" физиологов сверлила с другого конца тот же тоннель, что лингвистическая, скажем шире, палеопсихологическая мысль Марра. Но оба великих направления советской науки не завершили тогда этого встречного продвижения. И вот только теперь пробивается этот тоннель и воздух устремляется вдоль него.
Вернемся же к своему методу генетической трактовке морфологии речевых зон коры головного мозга и функциональных корковых нарушений речи. При этом мы остаемся на эволюционном этаже формирования мозга Homo sapiens и суггестивной стадии второй сигнальной системы.
Если интердикция (в начальном смысле, т. е. "интердикция I") еще чисто органический факт, хотя является аппаратом связи организмов, то суггестия на всем пути своего становления есть противодействие этой связи и новое преодоление этого противодействия и так далее. В этом качестве она совершенно специфична по отношению к собственной органической основе. Ключ ко всей истории второй сигнальной системы, движущая сила ее прогрессирующих трансформаций перемежающиеся реципрокные усилия воздействовать на поведение другого и противодействовать этому воздействию. Эта пружина, развертываясь, заставляла двигаться с этапа на этап развитие второй сигнальной системы, ибо ни на одной из противоположных друг другу побед невозможно было остановиться.
По первому разу интердикция могла быть отброшена, как мы помним, просто избеганием прямого контакта отселением, удалением. К числу первичных нейрофизиологических механизмов отбрасывания интердикции, судя по всему, следует отнести механизм персеверации (настаивания, многократного повторения). Он имеет довольно древние филогенетические корни в аппарате центральной нервной системы, наблюдается при некоторых нейродинамических состояниях у всех высших животных 55. Нельзя локализовать управление персеверацией у человека в каких-либо зонах коры головного мозга: как патологический симптом персеверация (непроизвольное "подражание себе") наблюдается при поражениях верхних слоев коры разных отделов, в частности, в лобной доле. Но кажется вероятным, что на подступах к возникновению второй сигнальной системы роль персеверации могла быть существенней. Инертное, самовоспроизводящееся "настаивание на своем" могло выгодно послужить как одной, так и противной стороне в отбрасывании или в утверждении и закреплении интердикции, следовательно, в генезе суггестии. Запомним, что последняя должна быть понята не просто как повеление, но как повеление, преодоление, преодолевающее отказ, впрочем, в противном случае оно даже и не повеление. Если же последующие исследования и не отведут специального места персеверации в филогенезе второй сигнальной системы, остается уверенность, что на позднейших этапах это довольно элементарное нервное устройство просыпалось снова и снова, становясь опорой всюду, где требовалось повторять, повторять, упорно повторять, в истории сознания, обобщения, ритуала, ритма.
Но отчетливое "отбрасывание" мы констатируем на уровне эхолалии. Правда, и этот феномен восходит к филогенетически древней основе непроизвольной имитации. Как патологический симптом при корковых поражениях у человека она называется эхопраксией, или эхокинезией. Однако применительно к речевой функции эта непроизвольная подражательная двигательная реакция, именуемая эхолалией, напоминает игру в теннис. Это повторение, но не своих слов, а чужих и в генезе, как говорилось выше и как доказывают обильные факты патологии и онтогенеза, повторение команд, прескрипций, требований. На требование "отдай" субъект отвечает словом "отдай" и тем освобождает себя от необходимости отдать. Иными словами, эхолалическая патология напоминает нам о той эпохе, когда в суггестии момент интердикции через имитацию еще доминировал над моментом конкретного дифференцированного предписания, что именно надлежит сделать. Эхолалическая реакция принимает данную определенную команду за команду вообще, безразличную к содержанию. Однако тут акцент уже не на застывании в моторике всего остального, а на факте общения: эхолалический "ответ" есть все-таки обмен словами, хотя и без обмена смыслами. Вся дальнейшая спираль развития речевого общения и будет перемежающимися уровнями обмена то обмена тождествами, то обмена нетождествами.
Если эхолалия обмен тождествами, то естественной защитой от нее являлась возникшая способность такой высокой фонологической дифференцировки звуков, которая при малейшем отклонении, "нарушении правил" приводит к фонологическому "непониманию" к неповторимости. В ответ на слово либо не последует ничего либо последует нечто не тождественное, тем самым "непонятное", нечто новое для первого партнера. Вот тут-то уже и завязываются наисложнейшие узлы второй сигнальной системы. Исследователь будет иметь дело с категориями "понимание" и "непонимание" последняя из них до уровня деформированной эхолалии навряд ли может быть применена, но дальше приобретает едва ли не доминирующее значение как психолингвистический феномен 56.
По словам психолингвиста Дж. А. Миллера, "нет психологического процесса более важного и в то же время более трудного для понимания, чем понимание, и нигде научная психология не разочаровывала в большей степени тех, кто обращался к ней за помощью" 57. В самом деле, даже с первого взгляда можно выделить противоречащие друг другу смыслы этого слова: а) собака "поняла" команду, если в точности ее выполнила; здесь нет предварительного психического понимания выполнение команды и есть понимание, т. е. стимул и реакция составляют единство; б) человек "понял" слова другого в смысле "распознал", "расслышал", "разобрал"; мы уже знаем, что в основе этого акта лежит повторение (громкое, внутреннее или редуцированное), т. е. идентификация слышимой и произносимой цепочек речевых звуков, повторение бессмысленное по своей сущности; этот акт "понимания" в чистом виде альтернативен по отношению к предыдущему: врач дает больному команду, а тот эхолалически повторяет ее вместо выполнения; в) идентификация не звуков речи, а содержания (смысла), т. е. новое повторение, но уже "другими словами": выявление инварианта, тождественности по существу двух лингвистически явно разных высказываний. Наконец, лишь бегло упомянем не касающийся нас здесь четвертый вариант: г) "понять" другого подчас значит идентифицировать скрытые мотивы его команды или высказывания и в зависимости от этого реагировать на его речь. Все это не только разные смыслы "понимания", но и альтернативные друг другу. Сейчас нам важна альтернативность "б" и "в".
Всякое средство отказа понимать (или быть понятым) можно называть средствами непонимания (или непонятности) , а соответствующий уровень эволюции уровнем непонимания (или непонятности). Хотя этот термин выглядит всего лишь негативным, так как конструируется с помощью приставки "не", он выражает позитивный феномен: не отсутствие понимания, а присутствие некоего обратного пониманию отношения и взаимодействия между людьми. Это есть общение посредством дезидентификации: посредством специального разрушения тождественности или сходства знаков. Точнее говоря, если идентификация, отождествление (сигнала с действием, фонемы с фонемой, названия с объектом, смысла со смыслом) служит каналом воздействия, то деструкция таких отождествлений или их запрещение служит преградой, барьером воздействию, что соответствует отношению недоступности, независимости. Чтобы возобновить воздействие, надо найти новый уровень и новый аппарат. Можно перечислить примерно такие этажи: 1) фонологический, 2) номинативный, 3) семантический, 4) синтаксическо-логический, 5) контекстуально-смысловой, 6) формально-символический 58. Однако все это продолжение тут нас не касается.
Фонологический этаж, он же эхолалические преодолевался становящимися людьми разными средствами. Так, сугубо физиологическим является факт наличия внизу коры головного мозга некоторых зон, искусственное возбуждение которых, не нарушая никаких прочих компонентов речевой функции, делает невозможным как раз повторение чужих слов (Н. А. Крышова). Видимо, природа пробовала создать такую самооборону, но ведь это было просто шагом вспять. Победили же эволюционные новации. О последних многое рассказывают нам те явления афазии, которые называются литеральными (буквенными) парафазиями: замена фонем противоположными, всяческие деструкции и декомпозиции звукового комплекса (слога, слова), в том числе инверсии и метатезы. Так, кстати, образовалась не только первичная бинарная оппозиция звуков, но и вся последующая множественность разных слов. Каждый раз это было антиэхолалией. Каждый раз новое слово было не только отличным и отличимым от другого, но как бы его опровержением, поэтому они уже не могли слиться обратно. Суть же дела состояла в том, что всякий ответ на слово таким преобразованным словом типа литеральной парафазии был одновременно и речеподражательным актом и, наоборот, актом не вполне речеподражательным, и эхолалией и неэхолалией отказом от эхолалической реакции и тем самым нейрофизиологическим прообразом ответа на вопрос или возражения на высказывание. Впрочем, только прообразом: не забудем, что речь идет о стадии, когда звукоиспускание было еще не связано со смыслами, а всего лишь тормозило нечто или высвобождало из-под торможения.
Из сказанного с необходимостью надлежит сделать вывод, что сама реакция эхолалического типа прошла две разные фазы: некогда она была самообороной от чьих-либо интердиктивных сигналов, но в дальнейшем сама превратилась в канал воздействия; видимо, даже чисто фонологическое "понимание" теперь стало вредным, или опасным, поэтому-то и пришлось изыскивать механизм, когда такое "понимание" хотя и есть (как голосовая подражательная реакция), но все же его одновременно и нет (ибо это деформированное подражание, наподобие передразнивания). В деталях переход от первой фазы ко второй неясен, но мы не рискуем ошибиться, сказав, что эхолалическая реакция стала сопровождаться какими-то ассоциируемыми с нею раздражениями и побуждениями в нервной системе. Следовательно, она тем самым эволюционировала навстречу собственно суггестии. Мы подойдем к этому факту с иной стороны в следующем разделе. Пока же отмечаем важный виток спирали "понимание непонимание": появление в акте эхолалии элементов действия "наоборот", т. е. подмена фонем противоположными по местоположению или по артикуляции создает очередной уровень "непонимания", или "нетождественного обмена".
Наконец, вот еще один механизм того же, восходящий, вероятно, к той же ранней поре к финалу чисто суггестивной стадии эволюции второй сигнальной системы. Это ответ молчанием.
Молчание может быть двоякого рода. Одно отвечает доречевому уровню. Это животное молчание. Другое перерыв, тормоз в речевом общении. Такое молчание второго рода было гигантским приобретением человечества. Оно тоже принадлежит к механизмам отказа от непосредственного выполнения суггестии, но и от парирования ее эхолалией или квазиэхолалией. Молчание генерализованное торможение речевой функции: тут уж нет подобия даже "неэквивалентного обмена", ибо в обмен не дается вообще ничего. Но это "ничего" весьма весомо. Во-первых, оно есть пауза разграничитель звуковых комплексов и тем самым фактор превращения неопределенно длительных звучаний в слова. Во-вторых, молчание в ответ на словесный раздражитель есть промежуточное звено к ответу действием, движением, но теперь предварительно пропущенным сквозь нейродинамическое сито дифференцирования словесных раздражителей. В-третьих, ответ молчанием есть первый шаг становления "внутреннего мира". Пока длится молчание, оно составляет оболочку для интериоризованных, внешне не проявляющихся реакций, будь то по речевому или неречевому типу. Следовательно, молчание это ворота к мышлению.
Но пока мы еще не вышли из мира суггестии. Мы только обозрели те барьеры, которые суггестия на этапе своей зрелости должна преодолевать, чтобы оставаться фактором принуждения в человеческом общении. Эти барьеры ее закалка. Суггестия вполне находит себя, когда она властна не над беззащитным, а над защищенным перечисленными средствами, т. е. преодолевает их.
Оставаясь еще в мире суггестии, мы тем самым исследуем только и исключительно систему материальных нейрофизиологических воздействий людей на поведение людей. Это поначалу просто своеобразное проявление тормозной доминанты, ее инверсия, вернее, целая серия инверсий в общении первобытнейших людей эпохи их отпочкования от троглодитов. На уровне суггестии вторая сигнальная система не имела никакого отношения к тому, что философия называет сознанием, как и познанием. Но она не только интериндивидуальный феномен, ибо все настойчивее затрагивала и то, что индивид делает в окружающей природной среде: сначала тормозила его действия, затем уже и требовала какого-то действия. Да и самые простые тормозящие команды, если они тормозят лишь определенное действие, ставят перед побуждаемым организмом немало задач конкретного осуществления: "иди сюда" или "пошел вон" могут требовать преодоления каких-либо препятствий, осуществления каких-либо предваряющих поступков; "отдай", "брось" могут потребовать отчленения или иных операций с предметами. Одним словом, если индивид не прибегает к попыткам не выполнить предписываемое, парируя суггестию, а подчиняется ему, то он оказывается перед вопросом: как его выполнить? Следовательно, чем более суггестия расчленяется, тем многообразнее и тоньше операционные задачи, возникающие перед человеком.
Мы помним, что суггестия по своему физиологическому генезису противостоит и противоречит первой сигнальной системе, а именно тому, что подсказывает и диктует организму его собственная сенсорная сфера. Теперь, с развитием суггестии, вся задняя надобласть коры мозга, включающая височную, теменную и затылочную области, должна приспосабливаться, пристраиваться к необходимости находить во внешней среде пути к выполнению заданий. Это требовало развития корковых анализаторов, развития перцептивной и ассоциативной систем особого, нового качества. Функции и органы гнозиса и праксиса приобрели у нас человеческую специфику вместе с развитием суггестии.
Таким образом, не тот "труд" каждого по отдельности, на который делает упор индивидуалистическая концепция антропогенеза, усовершенствовал мозг Homo sapiens, не та "деятельность" каждого одиночки перед лицом природы, а выполнение императивного задания, т. е. специфическое общение (суггестия). Другое дело, что тем самым суггестия несет в себе противоречие: зачинает согласование двух сигнальных систем, из противопоставления которых она изошла. Это противоречие окажется продуктивным: оно приведет к контрсуггестии. Однако это произойдет на более позднем этапе эволюции.
Здесь остается внести одно разъяснение к сказанному в настоящем разделе о суггестии. Могло создаться впечатление, что ранние неоантропы состояли из внушающих (суггесторов) и внушаемых (суггестентов); вторые то поддавались, то пытались противиться, то снова поддавались воздействующему влиянию (инфлюации) первых. Однако я просто рассматривал явление и его осложнения сами по себе, отвлекаясь от вопроса, кто именно состоял в данном отношении, т. е. всегда ли та же роль исполнялась той же особью. Теперь, дабы выпятить, что это была абстракция и в противовес возможному недоразумению, повторим противоположную модель: каждая особь играла то одну роль, то обратную и нимало не срасталась с ними. Но видимо, обе модели неистинны, во всяком случае есть еще одна, гораздо более интересная для исследователя.
Мы все время оперировали двумя партнерами, вернее, двумя сторонами (ибо каждый "партнер" мог быть и множественным). Представим себе теперь, что перед нами три действующих лица, т. е. три соучаствующих стороны. В таком случае инициатором или соучастником всякой "непонятности", всякого "барьера" может быть и сам суггестор, если он не намерен воздействовать на поведение некоторых реципиентов, именно тех, которые владеют "кодом" самозащиты, или же, напротив, намерен воздействовать только на них, минуя остальных. Кстати, мы тем самым возвращаем слову "код" его настоящее значение, утраченное современной кибернетикой: "код" может быть только укрытием чего-то от кого-то, т. е. необходимо подразумевает трех соучастников кодирующего, декодирующего и акодирующего (не владеющего кодом). В противном случае связь первых двух звеньев столь же бессмысленно величать "кодом", как величают "запоминающим устройством" депо или склад чего-либо.
Итак, метаморфозы суггестии, намеченные выше, вполне согласуются с такой антропогенетической канвой: три соучастника это три градации, которые мы выше наметили в неустойчивом переходном мире ранних (ископаемых) неоантропов, а именно: 1) еще весьма близкие к палеоантропам, т. е. полунеандерталоидный тип, 2) средний тип, 3) наиболее продвинутые в сторону сапиентации. Все вместе они, или по крайней мере второй и третий тип, стояли в биологическом противоречии, каковому противоречию и соответствует первоначальная завязь суггестии. Она достигает все большей зрелости внутри этого мира ранних неоантропов, причем наиболее элементарные формы суггестии действительны по отношению к более примитивному типу, а более сапиентные варианты неоантропов избегают воздействия суггестии благодаря вырабатывающимся предохранительным ограждениям. Чем более усложненный вариант суггестии мы рассматриваем, тем более он отвечает отношениям уже между сапиентными формами, становясь "непонятным" для отставших. Естественный отбор весьма энергично закреплял формирование соответствующих устройств (эхолалических, парафазических и др.) в мозге неоантропов и размывал средний тип; все дальше в стороне от эволюции суггестии оставался неандерталоидный тип. Полная зрелость суггестии отвечает завершению дивергенции. Но к этому времени среди самих Homo sapiens уже распространилось взаимное обособление общностей по принципу "кодирования" своей общности от чужих побуждений, т. е. самозащиты "непониманием" от повелений, действительных лишь среди соседей.
Дата добавления: 2015-04-15; просмотров: 1808;