БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕ .
Оценка параметра 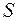 и ОП качества вскрытияпродуктивного пласта
и ОП качества вскрытияпродуктивного пласта
( 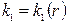 пласт неоднородный k = var)
пласт неоднородный k = var)
В том случае, когда приствольная зона скважины 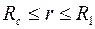 представляет собой область непрерывного изменения проницаемости
представляет собой область непрерывного изменения проницаемости 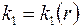 , уравнение неразрывности (3.57) видоизменится:
, уравнение неразрывности (3.57) видоизменится:
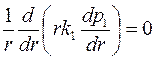 . .
| (3.72) |
Для удаленной части пласта 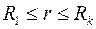 распределение давления
распределение давления 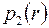 соответствует решению (3.51), а для приствольной зоны путем интегрирования (3.56) находим
соответствует решению (3.51), а для приствольной зоны путем интегрирования (3.56) находим
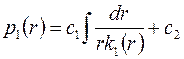 . .
| (3.73) |
Примем закономерность изменения проницаемости в области в виде
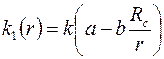 , ,
|
где 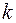 – проницаемость удаленной части пласта, т. е. при
– проницаемость удаленной части пласта, т. е. при
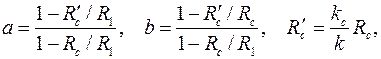
|
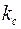 – проницаемость стенки скважины
– проницаемость стенки скважины 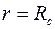 .
.
После подстановки 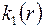 в (3.73),интегрирования и определения постоянных
в (3.73),интегрирования и определения постоянных 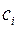 из граничных условий (3.68) получим следующее решение задачи:
из граничных условий (3.68) получим следующее решение задачи:
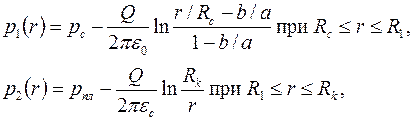
|
где 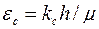 , а расход
, а расход 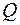 вычисляется по обобщенной формуле Дюпюи (3.65), в которой приведенный радиус скважины надо принять
вычисляется по обобщенной формуле Дюпюи (3.65), в которой приведенный радиус скважины надо принять
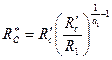 . .
|
Используя сходство этой формулы с формулой (3.70), легко найти
параметр , исходя непосредственно из формулы (3.71):
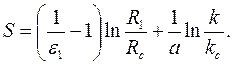
|
Пусть, например, при бурении проницаемого интервала 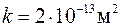
на стенке скважины сформирована глинистая корка проницаемостью 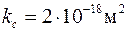 , т. е.
, т. е. 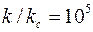 и
и 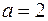 . Принимая
. Принимая 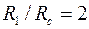 и
и 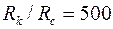 , получим
, получим
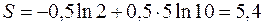 и
и 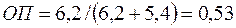 ,
,
т. е. поглощение фильтрата бурового раствора уменьшится более чем в 2 раза.
5. Плоская фильтрация в вертикально-трещиноватом пласте
Если пласт содержит упорядоченную систему трещин, то в нем благодаря анизотропии проницаемости плоско-радиальный характер фильтрации не будет иметь место (см. разд. 2).
Рассмотрим случай, когда одно из главных направлений анизотропии Ox3 совпадает с направлением оси скважины Oz (например, упорядоченная система вертикальных трещин в вертикальной скважине). Тогда два других главных направления анизотропии Ох1 и Ох2 расположены в плоскости 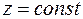 , т. е. параллельно кровле и подошве пласта. При заданных однородных граничных условиях в скважине и на поверхности питания (3.55) фильтрация будет плоской, так как
, т. е. параллельно кровле и подошве пласта. При заданных однородных граничных условиях в скважине и на поверхности питания (3.55) фильтрация будет плоской, так как 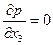 , но не радиальной. В плоскости х1х2 имеют место обобщенный закон Дарси [см. формулу (2.40)]
, но не радиальной. В плоскости х1х2 имеют место обобщенный закон Дарси [см. формулу (2.40)]
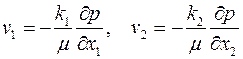 , ,
|
и соответствующее ему уравнение неразрывности [см. формулу (2.42)]
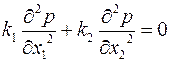 . .
| (3.74) |
Как было сказано в разд. 2, введением новой системы координат
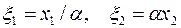
| (3.75) |
уравнение (3.74),заданное в анизотропной плоскости х1х2, преобразуется в уравнение Лапласа
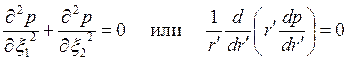 . .
| (3.76) |
для изотропной плоскости 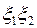 , проницаемость которой
, проницаемость которой
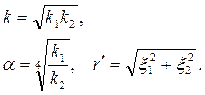
|
Принимая скважину в качестве источника (или стока) интенсивностью 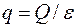 , получим, аналогично (3.62), поле давления
, получим, аналогично (3.62), поле давления
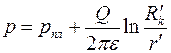 . .
| (3.77) |
где 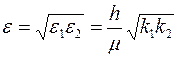 ,
, 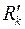 – радиус контура питания в плоскости . Отсюда следует, что эквипотенциальной поверхностью
– радиус контура питания в плоскости . Отсюда следует, что эквипотенциальной поверхностью 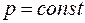 являются: окружность
являются: окружность 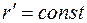 в плоскости и эллипс
в плоскости и эллипс 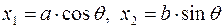 в плоскости х1х2, где
в плоскости х1х2, где 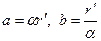 – полуоси эллипса.
– полуоси эллипса.
Это означает, что контуром питания (где 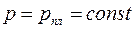 ) в анизотропном пласте может быть только эллипс
) в анизотропном пласте может быть только эллипс
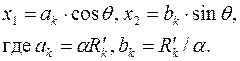
| (3.78) |
Согласно (3.59) этому эллипсу в плоскости соответствует окружность 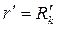 . В то же время окружность преобразуется в эллипс
. В то же время окружность преобразуется в эллипс
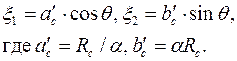
| (3.79) |
Поэтому в строгой постановке первая основная граничная задача формулируется так: найти решение уравнения (3.76), удовлетворяющее условию 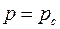 в точках эллипса (3.79) и условию
в точках эллипса (3.79) и условию 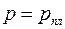 на окружности
на окружности 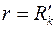 .
.
Однако для определения расхода ‚ достаточно хорошее приближение получается, если эллипс (3.79) заменить эквивалентной окружностью радиуса
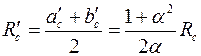 . .
| (3.80) |
Используя в (3.61) условие при 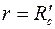 получим
получим
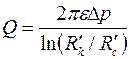 . .
| (3.81) |
Если истинный эллиптический контур питания (3.78) заменить условным – окружностью радиуса
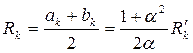
| (3.82) |
то, выразив через 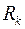 и подставив полученное выражение и соотношение (3.80) в (3.81) придем к обычной формуле Дюпюи (3.65), в которой
и подставив полученное выражение и соотношение (3.80) в (3.81) придем к обычной формуле Дюпюи (3.65), в которой 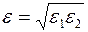 , а приведенный радиус скважины, приведенные коэффициенты гидропроводности и продуктивности надо принять равными:
, а приведенный радиус скважины, приведенные коэффициенты гидропроводности и продуктивности надо принять равными:
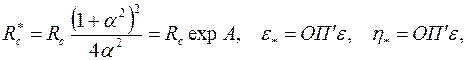
| (3.83) |
где 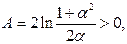
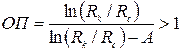 . .
| (3.84) |
Отсюда следует, что при прочих равных условиях в анизотропном пласте расход жидкости выше, чем в изотропном пласте эквивалентной гидропроводности .
В нижеследующей таблице приведены значения 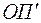 при нескольких параметрах анизотропии
при нескольких параметрах анизотропии 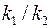 и
и 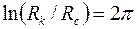 .
.
|
| 102 | 103 | 104 | |||
|
| 1,03 | 1,05 | 1,15 | 1,21 | 1,50 | 2,05 |
Видно, что влияние анизотропии заметно при больших отношениях 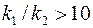 .
.
6. Определение расхода в неоднородном анизотропном пласте
Если после вскрытия пласта проницаемости 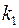 и
и 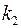 в приствольной зоне скважины изменились и стали равными
в приствольной зоне скважины изменились и стали равными 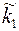 и
и 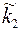 то возникает задача об определении расхода в неоднородном анизотропном пласте. Приближенное решение этой задачи может быть без труда найдено при следующих условиях:
то возникает задача об определении расхода в неоднородном анизотропном пласте. Приближенное решение этой задачи может быть без труда найдено при следующих условиях:
главные направления проницаемостей в приствольной зоне и удаленной части пласта совпадают;
границей раздела областями является эллипс
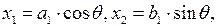
| (3.85) |
где 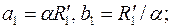
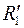 – радиус границы раздела в преобразованной плоскости .
– радиус границы раздела в преобразованной плоскости .
Обозначим давление на общей границе через 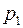 и рассмотрим каждую из областей независимо друг от друга.
и рассмотрим каждую из областей независимо друг от друга.
Так как подобным эллипсам (3.78) и (3.85) в плоскости соответствуют концентрические окружности 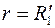 и , то для удаленной части пласта имеем [см. формулу (3.81)]
и , то для удаленной части пласта имеем [см. формулу (3.81)]
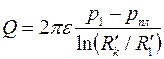 , ,
| (3.86) |
где –приведенная гидропроводность удаленной части пласта. Рассматривая приствольную зону скважины, замечаем, что здесь преобразование системы координат х1х2 в осуществляется с помощью другого параметра анизотропии 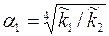 ,т. е.
,т. е.
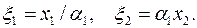
|
Следовательно, границы этой области: эллипс (3.69) и окружность преобразуются в эллипсы с соответствующими полуосями
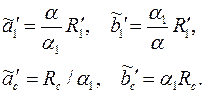
|
Заменив эти эллипсы эквивалентными окружностями, радиусы которых равны
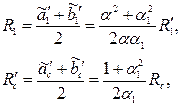
| (3.87) |
получим приближенную формулу для расхода жидкости
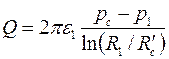 , ,
| (3.88) |
где 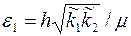 – приведенная гидропроводность призабойной части пласта.
– приведенная гидропроводность призабойной части пласта.
Определив из равенства правых частей (3.86) и (3.88), после преобразования получим следующую обобщенную формулу Дюпюи:
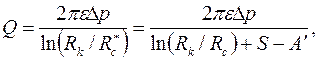 , ,
| (3.89) |
где
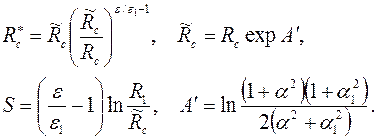 . .
|
Видно, что при 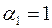 и
и 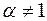 имеем
имеем 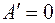 , т. е. влияние анизотропии исчезает, если призабойная зона скважины в результате кольматации приобрела свойства изотропной среды. Аналогичный результат имеет место при
, т. е. влияние анизотропии исчезает, если призабойная зона скважины в результате кольматации приобрела свойства изотропной среды. Аналогичный результат имеет место при 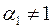 и
и 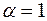 , что возможно, например, при гидроразрыве изотропного пласта. Отсюда следует вывод гидроразрыв гранулярного коллектора в ПЗ не может привести к заметному росту продуктивности скважины. Его положительная роль сводится к разрушению зоны кольматации и тем самым восстановлению потенциальной продуктивности пласта. Только при гидроразрыве анизотропного пласта, когда
, что возможно, например, при гидроразрыве изотропного пласта. Отсюда следует вывод гидроразрыв гранулярного коллектора в ПЗ не может привести к заметному росту продуктивности скважины. Его положительная роль сводится к разрушению зоны кольматации и тем самым восстановлению потенциальной продуктивности пласта. Только при гидроразрыве анизотропного пласта, когда 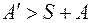 , продуктивность скважины может быть увеличена.
, продуктивность скважины может быть увеличена.
7. Несовершенное вскрытие пластов
Фильтрация, отличная от плоско-радиальной, возникает и в том случае, когда пласт вскрыт не на всю мощность, а частично или часть пласта перекрыта обсадной колонной, или связь пластовой и скважинной жидкостей осуществляется через перфорационные отверстия в колонне.
В этих случаях говорят о несовершенном вскрытии пласта и задают граничное условие лишь на открытой части поверхности , а на остальной условие непроницаемости 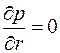 . Течение жидкости в таких условиях вблизи скважины пространственно, и, естественно, решение задачи фильтрации усложняется.
. Течение жидкости в таких условиях вблизи скважины пространственно, и, естественно, решение задачи фильтрации усложняется.
Известны различные приближенные аналитические решения этих задач и экспериментальные исследования на моделях, учитывающие тот или иной вид несовершенства вскрытия пласта.
Общий вывод, который следует из полученных решений, сводится к тому, что расход жидкости и в этих случаях вычисляется по обобщенной формуле Дюпюи (3.49), где приведенный радиус скважины
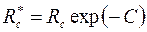 , ,
| (3.90) |
здесь 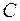 – показатель фильтрационного сопротивления, связанный с несовершенством вскрытия пласта.
– показатель фильтрационного сопротивления, связанный с несовершенством вскрытия пласта.
Отношение расхода жидкости при несовершенном вскрытии к расходу 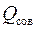 при совершенном вскрытии пласта в тех же условиях определяют аналогично параметру ОП [см. формулу (3.66)]
при совершенном вскрытии пласта в тех же условиях определяют аналогично параметру ОП [см. формулу (3.66)]
коэффициент сопротивления:
| (3.91) |
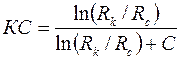 .
.
В общем случае 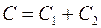 где
где 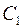 и
и 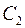 – показатели сопротивления, обусловленные несовершенством по степени и характеру вскрытия пласта. Для случая вскрытия части пласта
– показатели сопротивления, обусловленные несовершенством по степени и характеру вскрытия пласта. Для случая вскрытия части пласта 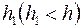 Маскет, используя метод источников, нашел, что при
Маскет, используя метод источников, нашел, что при 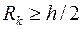 показатель несовершенства по степени вскрытия можно определить по формуле
показатель несовершенства по степени вскрытия можно определить по формуле
| (3.91) |
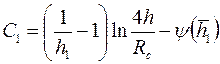 .
.
Здесь 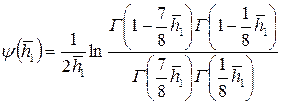 ,
,
где 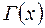 – гамма-функция (известная, табулированная функция);
– гамма-функция (известная, табулированная функция); 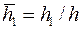 .
.
Представление о функции 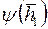 и показателе
и показателе 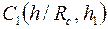 дает табл. 3.
дает табл. 3.
Таблица 3
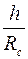
| 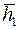
| |||||||
| 0,9 | 0,8 | 0,7 | 0,6 | 0,5 | 0,4 | 0,3 | 0,2 | |
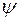
| ||||||||
| 0,43 | 0,84 | 1,38 | 2,04 | 2,93 | 4,33 | 7,1 | 13,11 | |
| 0,16 | 0,47 | 0,91 | 1,52 | 2,35 | 2,62 | 5,35 | 8,1 | |
| 0,24 | 0,65 | 1,21 | 1,98 | 3,04 | 3,65 | 6,87 | 10,87 | |
| 0,41 | 1,05 | 1,89 | 3,05 | 4,66 | 6,07 | 10,63 | 17,39 | |
| 0,49 | 1,22 | 2,19 | 3,52 | 5,35 | 7,11 | 12,24 | 20,08 |
Например, при Rc = 0,1 м, h = 20 м, h1 = 10 м, согласно таблице при h/Rc=200 и h1=0,5, получим С1=3,35, что при соответствует коэффициенту сопротивления КС = 0,65.
Существенное значение в этой задаче могут иметь различные проницаемости вдоль пласта 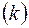 и в направлении, перпендикулярном к пласту
и в направлении, перпендикулярном к пласту 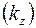 , т. е. анизотропия проницаемости. Доказано, что учесть этот фактор можно, если заменить истинную мощность пласта
, т. е. анизотропия проницаемости. Доказано, что учесть этот фактор можно, если заменить истинную мощность пласта 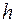 приведенной
приведенной 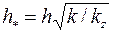 .
.
Если, например, 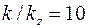 , то по данным предыдущего примера имеем
, то по данным предыдущего примера имеем 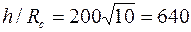 ,
, 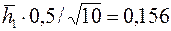 и, согласно формулам,
и, согласно формулам, 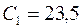 и
и 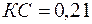 .
.
Несовершенство по характеру вскрытия имеет место, когда связь со скважиной осуществляется через круглые или щелевые отверстия в обсадной колонне. В этом случае показатель несовершенства может быть вычислен по следующим приближенным формулам:
| (3.93) |
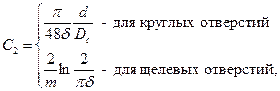
где 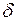 – открытая часть поверхности колонны;
– открытая часть поверхности колонны; 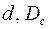 – диаметры перфорационных отверстий и скважины; т — число рядов щелей.
– диаметры перфорационных отверстий и скважины; т — число рядов щелей.
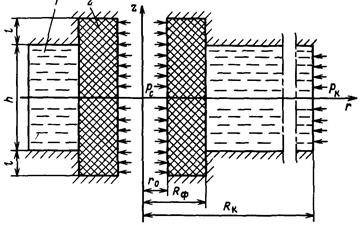
Рис. 3.5 Схема призабойной зоны скважины с искусственным фильтром
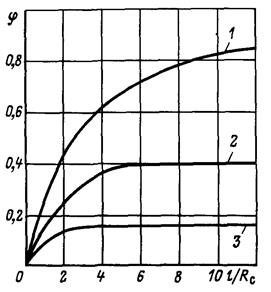
Рис. 3.6 Зависимость показателя снижения фильтрационного сопротивления от величины дополнительной зоны фильтрации при h/Re = 15: 1 2, 3 соответственно при Rф/Rc = 8; 5; 3.
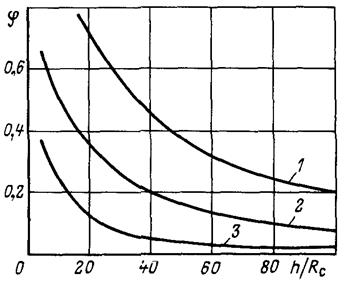
Рис. 3.7Зависимость показателя снижения фильтрационного сопротивления от мощности пласта и радиуса фильтра приl/Rф = 2: 1, 2, 3 соответственно
при Rф/Rc = 8; 5; 3
Приведем решение задачи, когда приствольная зона скважины оборудована искусственным фильтром (2)высотой 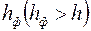 и проницаемостью
и проницаемостью 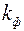 , отличной от проницаемости пласта (1)(рис. 3.5).
, отличной от проницаемости пласта (1)(рис. 3.5).
Приведенный радиус в этом случае
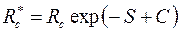 , ,
| (3.94) |
где – параметр «скин-эффекта» [см. формулу (3.71)]; 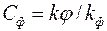 показатель снижения сопротивления, обусловленный наличием дополнительной зоны
показатель снижения сопротивления, обусловленный наличием дополнительной зоны 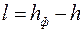 ; φ – функция безразмерных параметров
; φ – функция безразмерных параметров 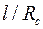 ,
, 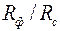 ,
, 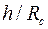 .
.
На рис. 3.6 показаны графики зависимости φ от при трех значениях отношения и 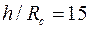 . Из него следует, что с увеличением функция
. Из него следует, что с увеличением функция 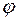 быстро растет до асимптотического значения, которое наступает при
быстро растет до асимптотического значения, которое наступает при 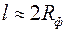 . Это доказывает нецелесообразность установки фильтра высотой больше чем
. Это доказывает нецелесообразность установки фильтра высотой больше чем 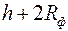 .
.
Влияние мощности пласта на φиллюстрируется графиками на рис.3.7 при тех же значениях и 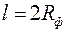 .
.
БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕ .
Как уже отмечалось, в процессе обмена веществ в организме происходят различные реакции окисления, которые объединяют термином биологическое окисление.
Основным типом биологического окисления является тканевое дыхание, протекающее в клеточных митохондриях (в связи с этим тканевое дыхание еще называют митохондриальным окислением)..
4.1. Тканевое дыхание.
Тканевое дыхание - это основной способ получения АТФ, используемый всеми клетками организма (кроме красных клеток крови).
В процессе тканевого дыхания от окисляемого вещества отнимаются два атома водорода (два протона и два электрона) и по дыхательной цепи, состоящей из ферментов и коферментов, передаются на молекулярный кислород - О2, доставляемый кровью из воздуха во все ткани организма. В результате присоединения атомов водорода к кислороду образуется вода. За счет энергии, выделяющейся при движении электронов по дыхательной цепи, в митохондриях осуществляется синтез АТФ из АДФ и фосфорной кислоты. Обычно образование одной молекулы воды сопровождается синтезом трех молекул АТФ.
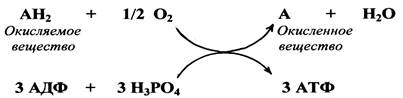 |
В упрощенном виде тканевое дыхание может быть представлено следующей схемой:
В качестве субстратов окисления (т.е. веществ, от которых отнимается водород) в тканевом дыхании используются разнообразные промежуточные продукты распада белков, углеводов и жиров. Однако, наиболее часто окислению подвергаются промежуточные продукты цикла трикарбоновых кислот (ЦТК) - цикла Кребса (изолимонная, a-кетоглутаровая, янтарная и яблочная кислоты). Цикл Кребса - это завершающий этап катаболизма, в ходе которого происходит окисление остатка уксусной кислоты, входящей в ацетилкофермент А, до СО2 и Н2О. В свою очередь, ацетилкофермент А - это универсальный метаболит организма, в который при своем распаде превращаются главные органические вещества - белки, углеводы и жиры (подробнее см. в главе 5 «Обмен углеводов»).
Тканевое дыхание представляет собой сложный ферментативный процесс. Все ферменты тканевого дыхания делятся на три группы: никотинамидные дегидрогеназы, флавиновые дегидрогеназы и цитохромы.
Никотинамидные дегидрогеназы отнимают два атома водорода от окисляемого субстрата и временно присоединяют их к своему коферменту НАД (никотинамид-аденин-динуклеотид). По строению НАД является динуклеотидом, в котором два нуклеодида соединяются между собой остатками фосфорной кислоты. В состав одного из нуклеодидов в качестве азотистого основания входит амид никотиновой кислоты (никитинамид, витамин РР), вторым нуклеотидом является аденозинмонофосфат (АМФ):
|
|
|
 |  | ||||
 |
Отнимаемые от окисляемого вещества атомы водорода присоединяются непосредственно к никотинамиду (отсюда название данных ферментов – никотинамидные дегидрогеназы), при этом НАД переходит в свою восстановленную форму НАД·Н2:
 АН2 + НАД А + НАД ·Н2
АН2 + НАД А + НАД ·Н2
Окисляемое Окисленное
вещество вещество
Флавиновые дегидрогеназы отщепляют два атома водорода от образовавшегося НАД·Н2 и временно присоединяют их к своему коферменту ФМН (флавин-мононуклеотид). По строению этот кофермент является мононуклеотидом, содержащим витамин В2 (рибофлавин):
|
|
|
 |  |





 Рибофлавин (витамин В2 )
Рибофлавин (витамин В2 )
Два атома водорода, отнимаемые флавиновыми дегидрогеназами от восстановленного НАД (НАД·Н2) присоединяются к флавину, выполняющего роль акцептора водорода. В результате этой стадии образуется восстановленная форма кофермента - ФМН·Н2:
 НАД ·Н2 + ФМН НАД + ФМН ·Н2
НАД ·Н2 + ФМН НАД + ФМН ·Н2
В некоторых случаях флавиновые дегидрогеназы, подобно никотинамидным, отнимают два атома водорода непосредственно от окисляемых веществ. Такие флавиновые дегидрогеназы используют кофермент ФАД (флавин-аденин-динуклеотид), похожий по строению на ФМН и тоже содержащий витамин В2:
 АН2 + ФАД А + ФАД ·Н2
АН2 + ФАД А + ФАД ·Н2
Окисляемое Окисленное
вещество вещество

 Следующая группа ферментов - цитохромы. Эти ферменты участвуют только в переносе электронов. По строению цитохромы похожи на одну из субъединиц гемоглобина. Молекула цитохрома состоит из полипептида и гема. Но в отличие от гемоглобина железо, входящее в гем цитохромов, имеет переменную валентность. Способность железа обратимо переходить из окисленной формы в восстановленную (Fe3+ + е Fe2+) обеспечивает возможность переноса электронов данными ферментами.
Следующая группа ферментов - цитохромы. Эти ферменты участвуют только в переносе электронов. По строению цитохромы похожи на одну из субъединиц гемоглобина. Молекула цитохрома состоит из полипептида и гема. Но в отличие от гемоглобина железо, входящее в гем цитохромов, имеет переменную валентность. Способность железа обратимо переходить из окисленной формы в восстановленную (Fe3+ + е Fe2+) обеспечивает возможность переноса электронов данными ферментами.
С помощью цитохромов (их имеется несколько: b, c, a, a3) электроны от восстановленных коферментов ФМН·Н2 и ФАД·Н2 передаются на молекулярный кислород (О2), который при этом переходит в активную, анионную форму - О2-. Далее, активный кислород (О2-) связывается с ионами водорода (протонами), которые тоже отщепляются от ФМН·Н2 или от ФАД·Н2. Присоединение ионов водорода к аниону кислорода приводит к образованию воды.
Таким образом, на всем протяжении дыхательной цепи наблюдается передвижение электронов. Движение электронов вызвано тем, что все участники дыхательной цепи располагаются по мере возрастания их окислительно-восстановительных потенциалов.
Окислительно-восстановительный потенциал или редокс-потенциал характеризует способность вещества принимать и удерживать электроны. Поэтому электроны переносятся от вещества с низким редокс-потенциалом к веществу с более высоким потенциалом.
Поскольку самое низкое значение редокс-потенциала имеет окисляемое вещество, а самое высокое – кислород, то в итоге электроны от окисляемого вещества поступают на молекулу кислорода.
Как уже отмечалось, движение электронов по дыхательной цепи сопровождается выделением энергии. Около половины энергии движения электронов аккумулируется в макроэргических связях молекул АТФ. Другая часть энергии выделяется в виде тепла. Синтез АТФ происходит при переносе электронов с НАД·Н2 на ФМН, с цитохрома b на цитохром с и с цитохрома a на цитохром a3. Всего при переносе двух атомов водорода на кислород (т.е. в расчете на одну образовавшуюся молекулу воды) синтезируется 3 молекулы АТФ.
Некоторые субстраты (жирные кислоты, янтарная кислота и др.) имеют более высокий редокс-потенциал, чем НАД. Поэтому они не могут окисляться никотинамидными дегидрогеназами. В этом случае отнятие атомов водорода от таких субстратов осуществляется флавиновыми дегидрогеназами. Из-за отсутствия никотинамидных дегидрогеназ при окислении таких веществ образуется только две молекулы АТФ.
На рис. 6 представлена схема тканевого дыхания, включающая все группы ферментов.
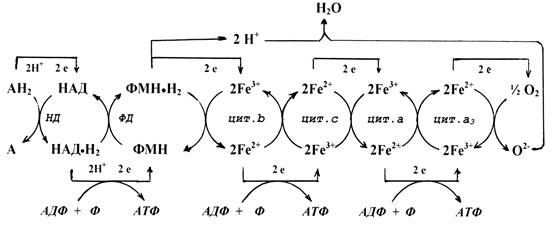
Рис. 6. Схема тканевого дыхания
Образование АТФ в процессе тканевого дыхания часто обозначается терминами: окислительное фосфорилирование, дыхательное фосфорилирование, аэробное фосфорилирование или аэробный синтез АТФ.
В сутки в организме за счет тканевого дыхания возникает не менее 40 кг АТФ, а у спортсменов еще больше. Поэтому этот процесс потребляет большое количество окисляемых веществ и кислорода.
При незначительной потребности клеток в АТФ тканевое дыхание протекает с низкой скоростью. Если клетка начинает использовать большое количество АТФ, то скорость тканевого дыхания возрастает и может достигнуть максимальных величин. Такой характер изменения скорости обусловлен тем, что активатором ферментов тканевого дыхания является избыток АДФ, который возникает в клетке только при интенсивном использовании АТФ.
Митохондрии, в которых протекает тканевое дыхание, имеются во всех клетках (кроме красных клеток крови) и представляют собою вытянутые палочковидные образования длиной 2-3 мкм и толщиной около 1 мкм. Количество митоходрий в клетках может достигать тысячи и более. Митохондрии снаружи окружены двойной мембраной. Внешняя мембрана гладкая, а внутренняя складчатая, с большой поверхностью. Ферменты тканевого дыхания встроены во внутреннюю мембрану и располагаются в ней в виде отдельных скоплений, называемых «дыхательными ансамблями» Каждый дыхательный ансамбль содержит все необходимые ферменты для обеспечения переноса электронов в процессе тканевого дыхания. Благодаря строго упорядоченному расположению ферментов в дыхательных ансамблях передвижение электронов по дыхательной цепи осуществляется с большой скоростью.
В клетках митохондрии часто располагаются в том месте, где используется энергия АТФ. В мышечных клетках митохондрии находятся около сократительных элементов – миофибрилл и обеспечивают энергией их сокращение в процессе мышечной работы. Под влиянием систематических тренировок количество митохондрий в мышечных клетках значительно увеличивается.
Как выше отмечалось, тканевое дыхание (митохондриальное окисление) является основным способом биологического окисления, т.е. окисления органических соединений в живом организме. Однако наряду с тканевым дыханием в организме еще имеются и другие способы окисления.
Дата добавления: 2015-03-07; просмотров: 2073;