Биотические факторы
Живое окружение организма составляет егобиотическую среду, а факторы этой среды называются биотическими (рис.1.2).
Внутривидовые взаимодействия между особями одного и того же вида заключаются во внутривидовой конкуренции и в групповом и массовом эффекте. В настоящее время эти эффекты называют также демографическими факторами, которые проявляются в основном в территориальном поведении животных (защита гнездовий, стадный образ жизни и т.д.).
Значительно более разнообразны межвидовые взаимоотношения. Влияние особей двух разных видов друг на друга может быть благоприятным, неблагоприятным или вообще никак не проявляться. Различные типы взаимоотношений рассмотрены в таблице 1.1.
| Таблица 1.1 | |||
| Типы межвидовых взаимоотношений | |||
| Тип взаимоотношения | Результат взаимодействия для видов | ||
| вид А | вид В | описание | |
| конкуренция | - | - | Конкуренция является в природе наиболее распространенным типом отношений, при котором две особи в борьбе за необходимые для жизни условия воздействуют друг на друга отрицательно. |
| нейтрализм | Тип отношений, при котором ни одна из популяций, обитающих на одной территории, не оказывает на другую никакого влияния. Например, белки и лоси. | ||
| комменсализм | + | Извлекая из хозяина (В) значительную пользу (пища, убежище), виды-комменсалы (А) не приносят ему никакой выгоды или заметного вреда. Например, многочисленные виды насекомых встречаются исключительно в муравейниках, норах грызунов, гнездах птиц, используя их как местообитание с более благоприятным микроклиматом. | |
| протокооперация | + | + | Обе популяции получают преимущества от совместного проживания, хотя их объединение не обязательно для выживания. Например, кишечнополостные защищают крабов своими стрекательными клетками, а взамен используют их как транспорт и получают от них кусочки утерянной крабами пищи. |
| мутуализм (симбиоз) | + | + | Длительное, неразделимое и взаимовыгодное отношение двух или более видов организмов. Например, жвачные животные переваривают клетчатку с помощью бактерий. Стоит только удалить этих симбионтов, и животные погибнут от голода. Классическим примером мутуализма является лишайник – симбиоз грибницы и водоросли. |
| аменсализм (антибиоз) | - | Один вид (А) причиняет вред другому (В), не извлекая при этом для себя никакой пользы. Чаще всего это те случаи, когда причиняемый вред заключается в изменении среды. Так поступает человек, разрушая и загрязняя окружающую среду. | |
| Продолжение таблицы 1.1 | |||
| паразитизм | + | - | Паразиты (А) питаются за счет другого организма, называемого хозяином (В). Паразиты живут на хозяине или внутри его организма на протяжении значительной части их жизненного цикла. Паразит использует для своей жизнедеятельности питательные вещества хозяина, тем самым постоянно ослабляя, а нередко убивая его. |
| хищничество | + | - | Особь одного вида (А), называемая хищником, питается организмами другого вида (В), называемого жертвой, причем хищник живет отдельно от жертвы. Например, птицы и насекомые. |
1.2. Экологические характеристики популяций
Популяция – совокупность особей одного вида, занимающих определенный ареал, свободно скрещивающихся друг с другом, имеющих общее происхождение, генетическую основу и в той или иной степени изолированных от других совокупностей данного вида.
Популяция характеризуется рядом показателей, которые разделяются на две группы – статистические и динамические.
Статические показатели характеризуют состояние популяции в определенный момент времени. К таким показателям относятся:
· численность – количество особей популяции в пределах ареала;
· плотность – число особей, приходящееся на единицу площади;
· половая структура – соотношение количества особей разного пола;
· возрастная структура – соотношение количества особей различного возраста.
Динамические показатели характеризуют процессы, протекающие в популяции за какой-то промежуток времени. Основными динамическими показателями являются:
· рождаемость – число рождающихся в популяции особей (∆Nb) за единицу времени (∆t). Мгновенная удельная рождаемость (b) – прирост численности особей популяции за единицу времени, отнесенный к начальной численности популяции (N), при ∆t→0:
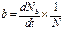 ;
;
· смертность – число погибших в популяции особей (∆Nd) за единицу времени(∆t). Мгновенная удельная смертность (b) – убыль численности особей популяции за единицу времени, отнесенный к начальной численности популяции (N), при ∆t→0;
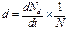 ;
;
· скорость роста популяции – скорость изменения численности популяции (увеличение или уменьшение) за ∆t, а при ∆t→0 ее можно определить как мгновенную скорость изменения численности популяции (r):
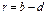 .
.
Если b = d, то r = 0, и популяция находится в стационарном состоянии.
Если популяция находится в благоприятных для нее условиях и значения всех экологических факторов близки к оптимуму, то численность популяции растет по закону геометрической прогрессии, размеры популяции при этом не стабилизируются. Т.е. имеет место экспоненциальный рост численности популяции (рис. 1.5а), который описывается J-образной кривой или уравнением: Nt=N0×ert, где: Nt – численность популяции в момент времени t; N0 – численность в начальный момент времени; e – основание натурального логарифма; r – скорость роста популяции. Особенно важно, что рост численности происходит в неизменных условиях внешней среды.
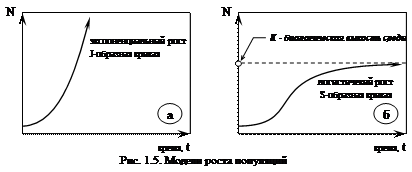
Однако реально такого не происходит, т.к. условия, сохраняющиеся длительное время постоянными, в природе не возможны. Обычно экспоненциальный рост наблюдается короткое время в начале развития популяции при наступлении благоприятных условий. Затем при смещении значений экологических факторов к пределам толерантности (например, недостаток элементов питания при увеличении плотности популяции) дальнейшее развитие популяции стабилизируется и описывается S-образной или логистической кривой роста (рис. 1.5б):
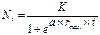 ,
,
где: rmax – максимальная скорость роста популяции в начальный период; а – константа; К – биологическая емкость среды в отношении особей данной популяции, т.е. число особей, которые могут нормально существовать в данной среде без заметного нарушения последней.
Однако плато на S‑образной кривой чаще всего не бывает гладким. Происходит постоянная флуктуация численности – колебания кривой около асимптоты К (рис. 1.6а). Флуктуации, вызванные как абиотическими, так и биотическими факторами, могут быть сезонными и годовыми. Регулярные флуктуации называют циклами (рис. 1.6б).
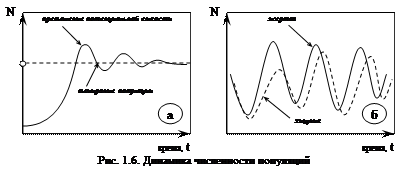
Классическим примером циклических колебаний являются колебания численности популяций хищника и жертвы. Так, циклы изменения численности популяций рыси и зайца составляют около 10 лет.
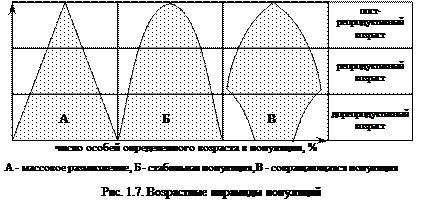
Часто для исследования возрастных структур популяций используют графические методы, например возрастные пирамиды популяции, широко используемые в демографических исследованиях (рис.1.7). Характер распределения особей популяции по возрастным группам позволяет прогнозировать динамику численности популяции на последующие периоды. Исследования численности популяций и определяющих ее факторов позволяет оценить последние и в отдельных случаях обеспечить ее биологическое pегулиpование.
Стремление организмов к выживанию и способы достижения этого называют экологической стратегией выживания. В уравнениях, описывающих рост популяции, были использованы параметры r и К. Виды, которые быстро размножаются, имеют высокое значение r и называются видами с r‑стратегией выживания. Стратегию видов с относительно низким значением r называют К-стратегией. Скорость размножения последних чувствительна к плотности популяции и остается близкой к уровню равновесия, определяемому величиной К. Некоторые характерные особенности r- и К-стратегий приведены в табл. 1.2.
| Таблица 1.2 | |
| Особенности экологических стратегий выживания | |
| r-виды | K-виды |
| Размножаются быстро (высокая плодовитость, короткое время регенерации) | Размножаются медленно (низкая плодовитость, большое время регенерации) |
| Скорость размножения мало зависит от плотности популяции | Скорость размножения зависит от плотности популяции (быстро увеличивается, если плотность падает) |
| Быстро расселяются на новые территории | Расселение медленное |
| Малые размеры особей | Относительно крупные особи |
| Слабо развиты защитные приспособления | Сильные защитные механизмы |
| Слабая конкурентоспособность | Сильные конкуренты |
| Примеры | |
| Бактерии, тли, однолетние растения | Кондор, альбатрос, кит, лев, человек |
1.3. Биотические сообщества
1.3.1. Понятие биоценоза, структура биоценоза
Совокупность видов живых организмов, обитающих совместно на определенной территории, составляет биотическое сообщество, при рассмотрении которого используют термин биоценоз.
Биоценоз (от гр. bios – жизнь, koinós – общий) – это совокупность растений, животных, микроорганизмов, населяющих участок суши или водоёма и характеризующихся определёнными отношениями как между собой, так и с абиотическими факторами среды. Более частными понятиями являются фитоценоз и зооценоз, представляющие собой соответственно совокупность растений или животных.
Биотоп (от гр. bios – жизнь, topos – место) – однородный по условиям жизни для определенных видов растений или животных или же для формирования определенного биоценоза участок территории. Таким образом, биотоп является местом жизни биоценоза.
Биоценоз представляет собой значительно более сложный уровень организации живого вещества, чем популяции, которые являются его составляющими. Биоценоз обладает сложной внутренней структурой. Обычно выделяют видовую и пространственную структуры биоценоза.
Видовая структура биоценоза характеризуется видовым разнообразием и количественным соотношением видов.
Видовое разнообразие – число видов живых организмов в данном сообществе или в данной области. Видовое разнообразие является важной качественной и количественной характеристикой устойчивости биотического сообщества. Различают α‑разнообразие (число видов в рассматриваемом биотопе), β‑разнообразие (число видов во всех биотопах данной области).
Количественное соотношение числа видов между собой показывает относительное число особей каждого вида, входящего в биоценоз. Так, например, соотношение 90:5:2:2:1 показывает, что среди ста особей в сообществе содержится 90 особей одного вида, 5 особей второго, по 2 особи третьего и четвертого и 1 особь пятого вида. Из этого соотношения видно, что первый вид является преобладающим, т.к. по числу особей составляет 90% всего сообщества.
Преобладающие по численности виды называют доминантными, или просто доминантами данного сообщества.
Эдификаторы – это виды, без которых сообщество не сможет существовать в том виде, в котором оно существует. Эти виды определяют микросреду (микроклимат) всего сообщества, и их удаление приведет к полному разрушению или трансформации биоценоза. Эдификаторами являются, как правило, растения (в лесу ель, сосна; в степи ковыль).
Внутри биоценозов существуют особые структурные объединения – консорции, представляющие собой группы разнородных организмов, поселяющихся на теле или в теле особи определенного вида – центрального члена консорции, – способного создавать вокруг себя определенную микросреду. Другие члены консорции могут формировать более мелкие консорции второго, третьего и т.д. порядка. Таким образом, можно сказать, что биоценоз является системой связанных между собой консорций.
Пространственная структура биоценоза представляет собой распределение видов или групп видов в пространстве биотопа в горизонтальном или вертикальном направлениях.
Особенно четко вертикальная пространственная структура прослеживается в фитоценозах лесов умеренного и тропического поясов. Такую структуру называют вертикальной ярусностью, или ярусным строением биоценоза. Например, в широколиственных лесах можно выделить пять ярусов: 1 – деревья первой величины (дуб, липа); 2 – деревья второй величины (рябина, яблоня, черемуха); 3 – кустарниковый подлесок (крушина, жимолость); 4 – высокотравье; 5, 6 – более низкие травы. Менее резко ярусность выражена и в травянистых сообществах. Она позволяет растениям более рационально использовать световой поток. В верхних ярусах преобладают светолюбивые растения, в средних – теневыносливые, в нижних – тенелюбивые.
Под воздействием растительности в различных ярусах изменяется микросреда. Меняется температурный градиент, газовый состав и другие абиотические факторы. Это способствует формированию ярусности фауны – от насекомых и птиц до млекопитающих.
Помимо ярусности, в биоценозе наблюдается горизонтальная пространственная структура – мозаичность, т.е изменение растительности и животного мира по горизонтали. Площадная мозаичность зависит от разнообразия видов, их соотношения, а также от ландшафтных и почвенных условий биотопа.
Наиболее благоприятные условия для существования множества видов характерны для переходных зон между сообществами – экотонами, а тенденцию к увеличению здесь видового разнообразия называют краевым эффектом. Классическим примером экотона является лесная опушка, на которой значительно богаче растительность, больше птиц, насекомых и других представителей фауны, чем в глубине леса. Этот эффект объясняется прежде всего тем, что в экотоны попадают виды из пограничных сообществ.
1.3.2. Экологическая ниша
Экологическая ниша – это место, занимаемое видом в природе, характеризующее его пространственное положение, функциональную роль в сообществе и отношение к абиотическим условиям существования. Важно то, что понятие экологической ниши не ограничивается физическим пространством, занятым видом, а наоборот основывается на экологических функциях вида в сообществе.
По мнению Ю.Одума, экологическую нишу можно образно представить как занятие или «профессию» организма в той системе видов, к которой он принадлежит, а его местообитание – как «адрес» вида. Характеристика экологической ниши дает знания о том, какие условия необходимы организму, чем он питается, чьей добычей он является и т.п.
Г.Хатчинсон предложил простую модель экологической ниши. Если на ортогональных проекциях отложить значения интенсивности воздействия различных факторов (f1, f2, …, fN), а из точек пределов толерантности определенного вида к этим факторам восстановить перпендикуляры, то ограниченное ими пространство будет соответствовать экологической нише данного вида (рис. 1.8).
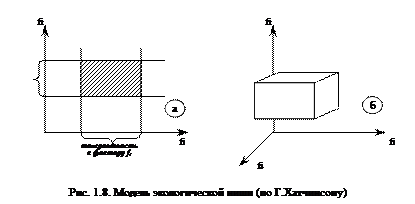
Таким образом, экологическая ниша представляет собой область комбинации таких значений факторов среды, в пределах которой данный вид может существовать неограниченно долго.
В случае действия на организм только двух факторов среды (температура, освещенность) можно говорить о двумерной нише (рис. 1.8а). Если учесть еще один фактор (влажность), то ниша будет трехмерной (рис. 1.8б). Однако в природной среде число экологических факторов, действующих на организмы определенного вида, очень велико, поэтому реальные экологические ниши всегда многомерны.
Фундаментальной экологической нишей называют нишу, определяемую только физиологическими потребностями организма. Она является «врожденной» нишей.
Реализованная экологическая ниша – это та часть фундаментальной ниши, которую данный вид или популяция смогли отстоять в конкурентной борьбе, т.е. ниша, в пределах которой вид существует в природе.
Конкурентное взаимодействие может касаться жизненного пространства, пищи или биогенных элементов, света, места укрытия и многих других жизненно важных факторов. Преимущества в конкурентной борьбе достигаются видами различными способами. При одинаковом доступе к ресурсу общего пользования один вид может иметь преимущество перед другим за счет более интенсивного размножения, потребления большего количества пищи или солнечной энергии, способности лучше защитить себя, адаптироваться к более широкому диапазону температур, освещенности или концентрации определенных вредных веществ.
В случае, когда экологические ниши двух видов перекрываются, возникают особо жесткие конкурентные взаимоотношения. Межвидовая конкуренция, независимо от того, что лежит в ее основе, может привести либо к установлению равновесия между двумя видами, либо к замене популяции одного вида популяцией другого, либо к тому, что один вид вытеснит другой в иное место или же заставит его перейти на использование иных ресурсов. Установлено, что два одинаковых в экологическом отношении и потребностях вида не могут существовать в одном месте, и рано или поздно один конкурент вытесняет другого. Это так называемый принцип конкурентного исключения, или принцип Гаузе.
Популяции некоторых видов живых организмов избегают конкуренции или снижают её переселением в другой регион с приемлемыми для себя условиями либо переходом на более труднодоступную или трудноусваиваемую пищу, либо сменой времени или места добычи корма. Так, например, ястребы питаются днем, совы – ночью; львы охотятся на более крупных животных, а леопарды – на более мелких; тенелюбивые растения не конкурируют со светолюбивыми. Для тропических лесов характерна сложившаяся стратификация животных и птиц по ярусам.
Процесс разделения популяциями видов пространства и ресурсов называется дифференциацией экологических ниш. Главный результат дифференциации – снижение конкуренции.
1.4. Экологические системы
Одним из основных понятий в экологии является понятие экосистемы. В целом под системойпонимается совокупность элементов, находящихся в отношениях и связях друг с другом, образующих определенную целостность, то есть структурно-функциональное единство.
Экологическая системапредставляет собой любую совокупность живых организмов и среды их обитания, взаимосвязанных обменом веществ, энергии, и информации, которую можно ограничить в пpостpанстве и во времени по значимым для конкретного исследования принципам. Ограничение может быть физико-химическим (например, граница капли воды, пруда, озера, острова) или связанным с круговоротом веществ, интенсивность которого внутри экосистемы выше, чем между экосистемой и внешним миром. Главной чертой понятия экосистемы является гибкость в отношении границ и масштаба. К экосистеме можно отнести как мелкий водоем, так и Мировой океан в целом.
Пример экосистемы – пруд с обитающими в нём растениями, рыбами, беспозвоночными животными, микроорганизмами, донными отложениями с характерными для него изменениями температуры, количеством растворённого в воде кислорода, составом воды и т.п., с определённой биологической продуктивностью; лес с лесной подстилкой, почвой, микроорганизмами, с населяющими его птицами, травоядными и хищными млекопитающими, с характерным для него распределением температуры и влажности воздуха, света, почвенных вод и др. факторов среды, с присущим ему обменом веществ и энергии.
Все экосистемы составляют иерархию в составе биосферы Земли и функционально связаны между собой. В связи с этим можно выделить различные структурные уровни в общей иерархии экосистем: микроэкосистемы (подушка лишайника и т.п.); мезоэкосистемы (озеро, степь, лесополоса и др.); макроэкосистемы (континент, океан); глобальная экосистема, или экосфера, – высший уровень интеграции всех экосистем мира.
Таким образом, устройство природы следует рассматривать как системное целое, состоящее из вложенных одна в другую экосистем, высшей из которых является уникальная глобальная экосистема – биосфера. В ее рамках происходит обмен энергией и веществом между всеми живыми и неживыми составляющими в масштабах планеты.
Главным предметом исследования при экосистемном подходе являются процессы трансформации вещества и энергии между живыми организмами и физической средой, т.е. биогеохимический круговорот веществ в экосистеме. Это позволяет дать интегрированную оценку результатов жизнедеятельности всей совокупности организмов, входящих в экосистему.
1.4.1. Свойства и структура экосистем
Прежде всего необходимо подчеркнуть, что экосистема является разновидностью всех систем природы, и поэтому практически все свойства, принципы и законы существования любых систем распространяются и на экосистемы.
Важным следствием иерархической организации экосистем является то, что, по мере объединения подсистем в более крупные, у новых систем возникают новые свойства, отсутствовавшие на предыдущем уровне. Такое наличие у системного целого особых свойств, не присущих его подсистемам и блокам, а также простой сумме элементов, называют эмерджентностью. Краткое определение эмерджентности звучит так: целое больше суммы его частей. Поэтому эмерджентные свойства экологической системы представляют собой не простой переход количества в качество, а являются особой формой интеграции, подчиняющейся иным законам формообразования, функционирования и эволюции. Такие качественно новые, эмерджентные свойства экологической единицы нельзя предсказать, исходя из свойств компонентов, составляющих эту единицу. Так, например, дерево или группу деревьев нельзя назвать лесом. Для существования леса необходимо не просто какое-то определенное количество деревьев, а сочетание всех его экологических компонентов (фито-, зоо- и микоценоз, микроклимат, круговорот веществ и т.д.), то есть эмерджентные свойства возникают в результате изменения функциональных связей этих компонентов, а не в результате изменения количества компонентов. Части не склеиваются, а интегрируются, обусловливая появление новых, до этого отсутствовавших свойств.
Очевидным является и тот факт, что никакая система не может сформироваться из абсолютно идентичных элементов. Закон необходимого разнообразия говорит о том, что любая система требует для своего существования минимально необходимого набора входящих в нее элементов. Количественно для каждого типа систем необходимое разнообразие различно. Нижний предел – не менее двух элементов, а верхний – бесконечность. Даже самые примитивные природные экосистемы состоят, как правило, из сотен, тысяч и более элементов. Например, трудно представить себе озеро, в котором присутствует только один-два вида растений и один вид рыб.
Согласно правилу конструктивной устойчивости надежная система может быть сложена из ненадежных элементов, или подсистем, не способных к самостоятельному существованию. По отношению к экосистемам это правило может быть уточнено следующим образом: устойчивая экологическая система может состоять из менее устойчивых компонентов, или подсистем; или – устойчивость экологической системы как единого целого всегда выше устойчивости каждого отдельного ее компонента, или подсистемы. Классическим примером тому могут служить лишайники, коралловые рифы, сообщества “социально организованных” насекомых. Один или несколько муравьев не способны выжить в естественных условиях, а колония муравьев, состоящая из большого числа схожих особей, может существовать долгое время.
Любая экологическая система существует в рамках определенных условий, выход за пределы которых может означать разрушение или значительное изменение параметров системы. Одним из главных законов функционирования систем является закон оптимальности, который гласит, что любая система функционирует с наибольшей эффективностью в некоторых характерных для нее пространственно-временных пределах (или: никакая система не может сужаться или расширяться бесконечно). Размер системы должен соответствовать выполняемым ею функциям, в противном случае она будет неэффективной или неконкурентноспособной.
Существующие природные системы являются открытыми системами: они обмениваются веществом, энергией и информацией с окружающим их миром. Очевидно, что в жизни экологических систем действуют общие законы сохранения и термодинамики, важные с точки зрения изучения потоков вещества и энергии. Масса и энергия подчиняются закону сохранения, то есть они не могут исчезать и появляться ни из чего. Закон сохранения массыв приложении к экосистемам звучит следующим образом: баланс вещества в системе количественно определяется разницей масс поступившего и вышедшего вещества за определенный промежуток времени. Большое значение в развитии экологических систем имеет закон максимизации энергии и информации: система всегда стремится к максимальному освоению поступающей к ней энергии и информации, что определяет ее устойчивость и конкурентоспособность.
Запасы вещества и энергии в экосистеме не безграничны, и они не могут бесконечно поддерживать функционирование экосистемы, поэтому необходим постоянный их приток извне. Не менее значительную роль играет круговорот веществ внутри самой экосистемы, который дает возможность использовать вещество многократно. Такой круговорот биогенных компонентов возможен только при наличии функционально различных групп организмов, способных осуществлять и поддерживать поток веществ, извлекаемых ими из окружающей среды. Процесс создания органического вещества в природе происходит одновременно с противоположным процессом – потреблением и разложением этого вещества вновь на исходные неорганические соединения. Совокупность этих процессов протекает в рамках экосистем различных уровней иерархии. Чтобы эти процессы были уравновешены, природа за миллиарды лет отработала определенную структуру живого вещества системы.
В природе можно выделить две основные группы организмов:
– автотрофы (“автос” – сам, “троф” – питаться, гр.) используют энергию неорганического происхождения для жизнедеятельности и синтеза органических веществ;
– гетеротрофы (“гетерос” – разный, гр.) используют энергию органических веществ для своей жизнедеятельности.
К автотрофам относятся продуценты (от лат. “продуцентис”- производящий), которые являются основой фоpмиpования и функциониpования экосистем. Это растения и микpооpганизмы, способные пpоизводить (пpодуциpовать) из неоpганического вещества оpганическое, используя энеpгию света или химические pеакции. Пpодуценты, использующие для пpодуциpования оpганического вещества солнечную энеpгию, называются гелиотpофами, а использующие химическую энеpгию – хемотpофами.
К гетеротрофам относятся консументы и редуценты.
Консументы (от лат. “консумо”– употреблять, расходовать)– организмы, потребляющие созданную продуцентами органику как источник вещества и энергии для своей жизнедеятельности. Консументы – это самые разнообразные организмы (от микроорганизмов до синих китов): простейшие, насекомые, пресмыкающиеся, рыбы, птицы и, наконец, млекопитающие, включая человека. Консументы, в свою очередь, подразделяются на ряд подгрупп в соответствии с различиями в источниках их питания. Животные, питающиеся непосредственно продуцентами, называются первичными консументами, или консументами первого порядка. Их самих употребляют в пищу вторичные консументы. Например, кролик – это консумент первого порядка, а лиса – консумент второго порядка. Первичные консументы, питающиеся только растениями, называются растительноядными, или фитофагами. Консументы второго и более высоких порядков – плотоядные, или зоофаги. Виды, употребляющие в пищу как растения, так и животных, относятся к всеядным.
Pедуценты (от лат. “редуцере”– уменьшение, упрощение строения)– оpганизмы, pазлагающие оpганические остатки (детpит) и экскpементы животных до минеpальных солей, котоpые возвpащаются чеpез почвенные pаствоpы обpатно коpням pастений. Редуценты замыкают биогеохимический круговорот веществ, разлагая органику на исходные неорганические составляющие. Редуценты в экосистемах представлены в основном микроорганизмами и грибами.
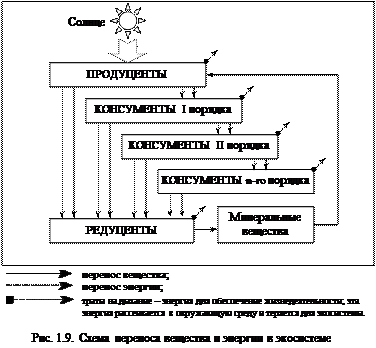
Таким образом, несмотря на многообразие экосистем, все они обладают структурнымсходством. В каждой из них можно выделить синтезирующие организмы – продуценты, различные уровни консументов, детритофагов и редуцентов. Они и составляют биотическую структуру экосистем, элементы которой объединены многочисленными связями, вещественными и энергетическими потоками(рис. 1.9).
1.4.2. Энергия экосистем. Экологические пирамиды
Энергию определяют как способность производить работу. Свойства энергии описываются законами термодинамики.
Первый закон термодинамики, или закон сохранения энергии, утверждает, что энергия может переходить из одной формы в другую, но она не исчезает и не создается заново. Второй закон термодинамики, или закон энтропии, утверждает, что в замкнутой системе энтропия (или мера беспорядка) может только возрастать. Применительно к энергии в экосистемах удобна следующая формулировка: процессы, связанные с превращениями энергии, могут происходить самопроизвольно только при условии, что энергия переходит из концентрированной формы в рассеянную, то есть деградирует.
У живых систем есть одно принципиальное отличие от неживых – они совершают постоянную работу против рассеивания энергии в окружающую среду. Жизнь – это единственный на Земле естественный самопроизвольный процесс, в котором энтропия системы уменьшается, т.е. упорядоченность системы возрастает. Такой процесс образования порядка в системе из хаоса окружающей среды называется самоорганизацией.
Таким образом, любая живая система, в том числе и экосистема, поддерживает свою жизнедеятельность благодаря, во-первых, наличию в окружающей среде в избытке солнечной энергии; во-вторых, способности за счет устройства составляющих ее компонентов эту энергию улавливать и концентрировать, а использовав, рассеивать в окружающую среду.
Основным источником энергии в экосистемах является солнечная энергия, которая “запасается” автотрофными растениями в энергию молекул органического вещества с помощью процессов фотосинтеза. Растения за счет содержащегося в них пигмента хлорофилла улавливают энергию излучения Солнца и используют ее для синтеза основы любого органического вещества – глюкозы C6H12O6:
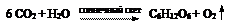
Излишек кислорода выделяется в атмосферу в газообразной форме. Энергия солнечного излучения преобразуется, таким образом, в потенциальную энергию, запасенную глюкозой. Всеми фотосинтезирующими организмами биосферы используется на процесс фотосинтеза около 1% всей солнечной энергии, дошедшей до поверхности Земли. Этого одного процента энергии достаточно для обеспечения ей всего живого вещества планеты и поддержания им состояния с низкой энтропией. Остальная часть энергии расходуется на нагрев атмосферы, суши и океана, переизлучается в космическое пространство в форме инфракрасного излучения.
Дыхание – это процесс разложения, обратный процессу фотосинтеза. При этом молекула глюкозы разлагается, а выделившаяся в результате разложения энергия расходуется в процессах синтеза других органических веществ. Из глюкозы вместе с получаемыми из почвы минеральными элементами питания – биогенами – образуются все ткани растительного мира – белки, углеводы, жиры, липиды, ДНК, РНК, то есть органическое вещество планеты.
Процессы синтеза и разложения органического вещества в экосистемах уравновешивают друг друга. Однако на уровне биосферы наблюдается некоторое отставание гетеротрофного разложения от автотрофного продуцирования во времени. Это является одним из важных свойств экосистем.
Перенос энергии пищи от ее источника – продуцента – через ряд организмов, происходящий путем поедания одних организмов другими, называется пищевой, или трофической цепью, т.е цепью взаимосвязанных видов, последовательно извлекающих органическое вещество и энергию из исходного пищевого вещества; каждое предыдущее звено является пищей для следующего.Трофические сети – сложные взаимоотношения в экологической системе, при которых разные компоненты потребляют разные объекты и сами служат пищей различным членам экосистемы. Упрощенная схема переноса вещества и энергии в экосистеме по трофическим сетям изображена на рисунке 1.9.
Пища, поглощаемая консументом, усваивается не полностью (15–20% у фитофагов, до 75% у зоофагов), т.е. от 25% до 85% запасенной на предыдущем трофическом уровне энергии теряется с экскрементами. Из усвоенной пищи значительная её часть расходуется на поддержание жизнедеятельности (на процессы метаболизма) организма, и только лишь небольшая часть идет на образование тканей, т.е на рост организма.
Таким образом, большая часть энергии при переходе с одного трофического уровня на другой, более высокий, теряется. Приблизительные потери составляют около 90%. Из количественных оценок, связанных с энергией, для трофических цепей известно правило десяти процентов: с одного трофического уровня на другой в среднем переходит около 10 % энергии, поступающей на предыдущий уровень.
Кроме органических веществ, играющих энергетическую и строительную роль, в круговорот веществ в экосистеме часто включаются химические компоненты, не специфичные для экосистем. Такими компонентами могут являться промышленные химикаты, ионы тяжелых металлов, яды и т.д. Эти вещества концентрируются, накапливаются в трофических цепях, т.е. происходит их биологическое накопление. Классическим примером эффекта биологического накопления является накопление ДДТ – ныне запрещенного инсектицида, применяемого ранее для борьбы с вредными насекомыми. Этим веществом опыляли комаров в дозах, не смертельных для других животных. Однако по мере накопления ДДТ, в заключительных звеньях трофической цепи (птицы) поглощаемая доза достигала высоких значений, что приводило к гибели птиц или неспособности воспроизводить потомство.
Любая экосистема характеризуется величиной продуктивности.
Продуктивность экологической системы – это скорость, с которой продуценты усваивают энергию, синтезируя органическое вещество, которое может быть использовано в качестве пищи. Различают первичную и вторичную продуктивность. Органическая масса, создаваемая продуцентами за единицу времени, называется первичной продукцией, а прирост за единицу времени массы консументов – вторичной продукцией.
В первичной продуктивности различают валовую и чистую продуктивность. Валовая первичная продуктивность – это скорость, с которой растения накапливают химическую энергию при фотосинтезе. Часть ее они тратят на поддержание собственной жизнедеятельности, эта энергия затем в виде теплоты выделяется в окружающую среду и теряется для экосистемы. Скорость накопления органического вещества продуцентами за вычетом расхода на собственную жизнедеятельность называется чистой первичной продуктивностью (она представляет величину прироста растения). Это энергия, которую могут использовать организмы следующих трофических уровней.
Все биотические компоненты экосистемы (продуценты, консументы и редуценты) составляют общую биомассу сообщества, которая может быть выражена как через весовые характеристики (килограммы, тонны), так и через энергетические (джоули, калории и т.п.).
Трофическую структуры  экосистем можно изобразить графически в виде экологических пирамид. Основанием пирамиды служит первый трофический уровень – уровень продуцентов, а последующие уровни образуют следующие этажи пирамиды. При этом высота всех блоков-этажей одинакова, а длина пропорциональна определенному показателю на соответствующем уровне.
экосистем можно изобразить графически в виде экологических пирамид. Основанием пирамиды служит первый трофический уровень – уровень продуцентов, а последующие уровни образуют следующие этажи пирамиды. При этом высота всех блоков-этажей одинакова, а длина пропорциональна определенному показателю на соответствующем уровне.
Известны три основных типа экологических пирамид:
1. пирамида чисел – отражает численность организмов на каждом уровне;
2. пирамида биомасс – показывает массу живого вещества;
3. пирамида энергии – характеризует изменение энергии (или первичной продукции) на последовательных трофических уровнях.
Пирамиды чисел и биомасс отражают статику системы, то есть характеризуют количество или биомассу организмов в определенный промежуток времени. Они не дают полной информации о трофической структуре экосистемы, хотя также позволяют решать ряд практических задач, особенно связанных с сохранением устойчивости экосистем.
В наземных экосистемах действует правило пирамиды биомасс: суммарная масса растений превышает массу всех травоядных, а их масса превышает биомассу хищников (рис. 1.11а). Для океанических экосистем данное правило не действительно, т.к. пирамида в данном случае имеет инверсный вид (рис. 1.11б), что говорит о тенденции накопления биомассы на более высоких уровнях.
Пирамида энергий является наилучшим графическим изображением трофической структуры экосистемы, поскольку она отражает динамику системы, то есть скорость прохождения энергии через пищевую цепь. Правило пирамиды энергии (или продукции) гласит: на каждом предыдущем трофическом уровне количество аккумулированной энергии (или биомассы), создаваемой за единицу времени, больше, чем на последующем. Пирамида энергий имеет универсальный характер и имеет сходный вид как в наземных, так и в океанических экосистемах.

Таким образом, для обеспечения энергией всех особей сообщества живых организмов экосистемы необходимо определенное количественное соотношение между продуцентами, консументами разных порядков и редуцентами. Для описания поведения энергии в экосистемах употребляют термин поток энергии, поскольку в отличие от циклического движения веществ,превращения энергии идутв одном направлении. Энергия, однажды использованная каким-либо организмом, превращается в тепло и утрачивается для экосистемы. Для своей жизнедеятельности каждый живой компонент, будь то организм или экосистема, должен получать от окружающей среды на входе постоянный приток дополнительной энергии. Живые замкнутые термодинамические системы невозможны.
Однако для жизнедеятельности любых организмов, а значит и системы в целом, только энергии недостаточно, они обязательно должны получать различные минеральные компоненты, микроэлементы, органические вещества, необходимые для построения молекул живого вещества.
1.4.3. Динамика экосистем
Для экосистем характерны те же динамические процессы, что и популяциям, сообществам, входящим в нее. Экосистемы испытывают цикличность функционирования, смену биоценозов и др.
Цикличность экосистем является, как правило, следствием суточных, сезонных и многолетних колебаний условий окружающей среды, а также эндогенных ритмов организмов и популяций. В районах с выраженным континентальным климатом, при большой разнице дневных и ночных температур наиболее четко проявляется суточная и сезонная цикличность, связанные, соответственно, с суточными и сезонными ритмами температуры, влажности и освещенности. Проявления этих видов цикличности можно легко наблюдать в условиях средней полосы России: ночной образ жизни хищников, впадение в спячку некоторых животных, дефолиация у деревьев (опадение листвы) и т.д. Менее выражена сезонная цикличность в районах с мягким муссонным климатом. Многолетняя цикличность может быть вызвана флуктуациями климата, такими, как неравномерное выпадение осадков по годам, периодическое повторение засух. Кроме того, многолетняя цикличность может быть связана с особенностями развития растений-эдификаторов.
Последовательная смена биоценозов, преемственно возникающая на одной и той же территории под воздействием природных факторов или воздействия человека, называется сукцессией (от лат. “сукцессио” – преемственность). Например, после лесного пожара горельник в течение многих лет заселяется сначала травами, потом кустарником, затем лиственными деревьями и в конечном итоге хвойным лесом. При этом последовательные сообщества, сменяющие друг друга, называются сериями, или стадиями. Конечным результатом сукцессии будет состояние стабилизированной экосистемы – климакс (климакс – лестница, "зрелая ступень", гр.). На этой стадии в экосистеме существует максимальное количество симбиотических связей, а на единицу энергии приходится максимальная биомасса.
Сукцессионные процессы в экосистемах инициируются, главным образом, изменением условий существования, которые, в свою очередь, могут быть вызваны как внешними воздействиями на систему (пожар, загрязнение и др.), так и нарастанием внутренних противоречий в системе (изменение почвы, смена доминантных видов и др.). При этом происходит нарушение существующего круговорота веществ и экологических связей между видами. Часть популяций, не способных выдержать конкуренции в изменившихся условиях среды, вытесняются другими популяциями, для которых эти условия являются более благоприятными; устойчивое состояние экосистемы (гомеостаз) на какое-то время восстанавливается.
По характеру и условиям возникновения выделяют два вида экологических сукцессий: первичные и вторичные.
Первичная сукцессия – это последовательное формирование и развитие сообществ на первоначально свободном субстрате, на участках, лишенных почв. Такими участками могут быть скалы, глины после оползня или обвала, остывшая лава, участки открытой добычи полезных ископаемых, с которых снят верхний слой почвы, и т.п. На таких бесплодных участках первичная сукцессия от голой скальной породы до зрелого леса может занять до 1000 лет. Первыми на такие участки заносятся споры, семена и вегетативные органы растений; происходит постепенная миграция животных с соседних территорий. Эта стадия называется стадией первых поселенцев. По мере формирования биоценоза происходит изменение экологических факторов участка: изменяется микроклимат, почва, видовой состав флоры и фауны. На начальных стадиях первичной сукцессии преобладающую роль играют r-виды, т.е. быстро размножающиеся виды, не требовательные к условиям среды. Виды с К-стратегией более конкурентоспособны, и в конце концов они вытесняют r-виды, которые тем временем перемещаются в другие нарушенные местообитания.
Примером первичной сукцессии является развитие на свободной территории ельника, который является последней стадией развития экосистем в климатических условиях Севера. Прорастая на свободных участках в одно и то же время, через 6–10 лет ель достигает 50–60 см, а береза 8–10 м. Т.е. сначала развиваются березняки, осинники, ольховники, под пологом которых растут ели. Под сомкнутыми кронами берез возникает свой микроклимат, формируются почвы, и ель продолжает расти в благоприятных для неё условиях. В итоге ель вырастает и формирует верхний ярус биоценоза, а новые проростки березы уже не способны расти в изменившихся условиях. Береза не выдерживает конкуренции за пространство и свет и вытесняется елью.
Более распространенным типом сукцессий является вторичная сукцессия – последовательное развитие сообщества на территории, в пределах которой естественная растительность была устранена или сильно нарушена, но почва не была уничтожена. Участками, на которых может наблюдаться вторичная сукцессия, могут быть заброшенные сельхозугодья, сгоревшие или вырубленные лесные участки, земли, где растительность уничтожена в результате ведения горностроительных работ, сильно загрязненные водные источники, затопленные ранее территории и т.п. Поскольку имеется в наличии какой-то слой почвы, новая растительность может появиться в течение всего нескольких недель. Вторичная сукцессия заканчивается стабильной стадией, как правило, через 150–250 лет.
Последовательный ряд постепенно и закономерно сменяющих друг друга в сукцессии сообществ называется сукцессионной серией. Она наблюдается в лесах, озерах, болотах, на стволах упавших деревьев и в пнях, в лужах и прудах и т.д. То есть сукцессии разномасштабны и иерархичны, так же как и сами экосистемы.
С энергетических позиций сукцессия – это такое неустойчивое состояние сообщества, которое характеризуется несоответствием двух показателей: валовой продуктивности и энергетических затрат всей системы на поддержание жизнедеятельности – дыхания.
В ходе сукцессии общая биомасса сообщества сначала возрастает, а продуктивность сообщества относительно высока, но затем темпы этого роста снижаются и на стадии климакса биомасса системы стабилизируется. В зрелых, устойчивых экосистемах практически весь прирост растительности поступает и расходуется в цепях питания гетеротрофами, поэтому чистая продуктивность сообщества приближается к нулю.
Значение этой закономерности важно в практической деятельности человека. Изымая избыток чистой продукции из сообществ, находящихся в начале развития сукцессии (напр., агроценозов), мы не подрываем основу существования сообщества. Вмешательство же в стабильные, климаксовые системы неминуемо вызывает нарушение сложившегося равновесия.
Таким образом, сообщество не может одновременно сочетать два противоположных свойства: быть высокостабильным и давать большой запас чистой продукции, который можно было бы изымать без вреда для него самого. Все искусственно создаваемые в сельскохозяйственной практике экосистемы полей, садов, пастбищных лугов, огородов, теплиц, то есть все агроценозы, представляют собой системы, специально поддерживаемые человеком на начальных стадиях сукцессии. В агроценозах используется именно их свойство производить высокую чистую продукцию, так как все конкурентные воздействия на культивируемые растения со стороны сорняков сдерживаются агротехническими мероприятиями, а формирование пищевых цепей за счет вредителей пресекается с помощью различных мер, например химической и биологической борьбы. Но эти сообщества неустойчивы, не способны к самовосстановлению и саморегулированию, подвержены угрозе гибели от массового размножения вредителей или болезней. Для их поддержания необходима постоянная деятельность людей.
2. БИОСФЕРА И ЧЕЛОВЕК
Биосфера (bios – жизнь, sphaira – сфера, шар, гр.) – сложная наружная оболочка Земли, населенная организмами, составляющими в совокупности живое вещество планеты. Это одна из важнейших сфер Земли, являющаяся основным компонентом природной среды, окружающей человека.
Понятие биосферы имеет два аспекта: с одной стороны, как специфическая оболочка земного пространства, с другой – как глобальная экосистема. С точки зрения составляющих компонентов биосфера – это нижняя часть атмосферы, гидросфера и верхняя часть литосферы, населенные живыми организмами, или, по выражению В.И. Вернадского, "область распространения живого вещества".
Все экологические ниши, пригодные для жизни, заняты биосферой, возникшей одновременно с появлением жизни на Земле (ок. 4 млрд лет назад) в виде примитивных протобиоценозов в первичном Мировом океане. Ок. 450 млн лет назад живые организмы стали заселять сушу, где их эволюция (возможно, в силу более жестких, чем в океане, экологических условий) ускорилась, и в результате соотношение числа видов животных и растений в Мировом океане и на суше составляет примерно 1:5. Основными факторами эволюции биосферы являются: абиотические (геологические, космические), биотические (изменчивость, т.е. мутации, наследственность, борьба за существование, естественный отбор), а также антропогенные, благодаря которым биосфера постепенно обретает черты ноосферы.
Таким образом, формирование и развитие биосферы неразрывно связано с эволюцией каждой из географических оболочек Земли.
Атмосфера ("атмос" – пар, гр.) – газовая оболочка нашей планеты, состоящая из смеси различных газов, водяных паров и пыли, граничащая с космическим пространством. Общая масса атмосферы – 5,15 • 1015т. Через атмосферу осуществляется обмен вещества Земли с космосом. Земля получает космическую пыль и метеоритный материал, теряет самые легкие газы: водород и гелий. Атмосфера Земли насквозь пронизывается радиацией Солнца, которая определяет тепловой режим поверхности планеты, вызывает диссоциацию молекул атмосферных газов и ионизацию атомов. На высоте от 10 до 50 км расположен слой озона, защищающий Землю от чрезмерного ультрафиолетового облучения. Обширная область разряженной верхней атмосферы состоит преимущественно из ионов. Эта область обозначается как ионосфера. Большая часть массы атмосферы имеет относительно однородный азотно-кислородный состав (азот – 75,5%, кислород – 23,14% от массы всей атмосферы). В тропосфере во взвешенном состоянии присутствуют также твердые и жидкие частицы, которые обычно называют аэрозолями. Атмосфера физически, химически и механически воздействует на литосферу и гидросферу, регулируя распределение тепла, влаги и различных веществ.
Гидросфера ("гидора" – вода, гр.) – водная оболочка Земли. По отношению к объему земного шара объем гидросферы не превышает 0,13%. На долю Мирового океана приходится 96,53% от общих мировых запасов воды; ледники – 1,74; подземные воды – 1,69; подземные льды в зоне вечной мерзлоты – 0,023, озера – 0,014, почвенная влага – 0,01, пары атмосферы – 0,01, болота – 0,0007, реки – 0,0002%.
Гидросфера находится в тесной взаимосвязи с литосферой (подземные воды), атмосферой (парообразная влага) и живым веществом биосферы, в которое она входит в качестве обязательного компонента. Вода способна растворять в себе многие вещества, поэтому любые воды гидросферы можно рассматривать в качестве естественных растворов различной степени концентрации. Даже наиболее чистые атмосферные воды содержат 10 – 50 мг/л растворенных веществ.
Мировой океан представляет собой уникальную природную систему. В ней происходит грандиозный процесс обмена и трансформации энергии и вещества нашей планеты. При этом различные физические, химические и биологические процессы объединяются, образуя единую природу океана – древнейшую область биосферы Земли. Со времени образования океана протекало изменение его природы под воздействием различных природных процессов: солнечного излучения, геологических и геохимических факторов и, что особенно важно, под влиянием биологических процессов.
В настоящее время можно считать, что в морской воде присутствуют все химические элементы таблицы Менделеева. Однако преобладающая часть растворенных веществ представлена немногими химическими элементами: O, H, Na, Mg, Ca, Cl, S. Некоторые из элементов, несмотря на относительно низкую концентрацию, играют важную роль в химических процессах моря и в морских организмах. В этом отношении ведущая роль принадлежит азоту, фосфору и кремнию, которые усваиваются живыми организмами, и их концентрация в морской воде контролируется ростом и размножением морских животных и растений.
Необходимо отметить одну особенность океанической воды – основные ионы, составляющие живое вещество, характеризуются постоянным соотношением во всем объеме Мирового океана. Это указывает на устойчивость динамического равновесия между количеством растворенных веществ, поступающих с поверхности континентов в океан, и их осаждением.
Литосфера – это верхняя твердая оболочка Земли, имеющая большую прочность и переходящая в нижележащую астеносферу, прочность которой относительно мала. Она включает земную кору и верхнюю мантию до глубин примерно 100–200 км. Выделяются два основных типа земной коры – континентальный и океанический. Между ними находится промежуточный тип, который называют субконтинентальным. Земная кора сложена породами различного типа и различного происхождения. Из них на осадочные породы приходится 9,2 %, на метаморфические – 20 % и на магматические – 70,8 %.
Общий химический состав земной коры определяют немногие элементы: O, Si, Al, Fe, Ca, Mg, Na, K, которые слагают основную ее массу. При этом наиболее распространенным элементом является кислород, составляющий едва ли не половину массы земной коры (>47,3 %). Он прочно связан химически с другими элементами в главных породообразующих минералах.
2.1. Учение о биосфере
Автором термина "биосфера" является французский естествоиспытатель Жан Батист Ламарк, который употребил его в 1803 г. в труде по гидрогеологии Франции для обозначения совокупности организмов, обитающих на земном шаре. Затем термин был забыт. В 1875 г. его "воскресил" профессор Венского университета геолог Эдуард Зюсс (1831 – 1914) в работе о строении Альп. Он ввел в науку представление о биосфере как особой оболочке земной коры, охваченной жизнью. В таком общем смысле впервые в 1914 г. использовал этот термин и В. И. Вернадский в статье об истории рубидия в земной коре.
Общее учение о биосфере создано в 20–30-х гг. XX в. В.И. Вернадским, развившем идеи В.В. Докучаева о комплексном естественно-историческом анализе взаимодействующих в природе разнокачественных объектов и явлений (факторов почвообразования) и выявлении самостоятельных природных объектов гетерогенной структуры и состава (почвы, природные зоны). Позднее идеи В.И. Вернадского сыграли ключевую роль в формировании мировоззрения современного человека, в понимании им своего места в природе и ответственности за будущее биосферы, в формировании новой экологической морали.
Человечество как живое вещество неразрывно связано с материально-энергетическими процессами определенной геологической оболочки Земли – ее биосферой. Оно не может физически быть от нее не зависимым ни на одну минуту. Однако и само человечество есть закономерный продукт развития биосферы, развития образующего ее живого вещества как природного тела.
2.1.1. Состав и структура биосферы
Живое вещество биосферы в общем занимает ничтожное пространство в масштабе всего земного шара. В.И. Вернадский показал, что все живые организмы Земли образуют единое целое – живое вещество планеты. Жизнь на Земле – самый выдающийся процесс на ее поверхности, получающий энергию Солнца и приводящий в движение большинство химических элементов Земли. Биосфера есть часть земного пространства, охваченного жизнью с ее активным химическим проявлением.
Биосфера включает не только область жизни (биогеосферу, фитогеосферу, витасферу), но и другие структуры Земли, генетически связанные с живым веществом. По Вернадскому, вещество биосферы состоит из семи разнообразных, но геологически взаимосвязанных частей: живое вещество; биогенное вещество (уголь, нефть, известняки); косное вещество (магматические породы); биокосное вещество (создается с помощью живых организмов); радиоактивное вещество; рассеянные атомы; вещество космического происхождения. В пределах биосферы везде встречается либо живое вещество, либо следы его биогеохимической деятельности. Газы атмосферы (кислород, азот, углекислота), природные воды, равно как и каустобиолиты (нефти, угли), известняки, глины и их метаморфические производные (сланцы, мраморы, граниты и др.) в своей основе созданы живым веществом планеты. Слои земной коры, лишённые в настоящее время живого вещества, но переработанные им в геологическом прошлом, Вернадский относил к области "былых биосфер".
Биосфера, являясь глобальной экосистемой (экосферой), как и любая экосистема, состоит из абиотической и биотической частей.
Абиотическая часть представлена:
– почвой и подстилающими ее породами, где в них обитают живые организмы, вступающие в обмен с веществом этих пород;
– атмосферным воздухом до высот, где возможны проявления жизни;
– водной средой океанов, рек и др. водоемов.
Биотическая часть состоит из всей совокупности живых организмов, которые в процессе своей жизнедеятельности, взаимодействуя с абиотической частью, осуществляют биогенный ток атомов.
В биосфере возможно существование организмов в любых возможных концентрациях – от единичных бактерий и спор в 1 см3 атмосферного воздуха и следов жизни в глубинах Мирового океана до мощных тропических лесов экваториальной зоны. По своим требованиям к условиям внешней среды организмы расселяются в разных верхних горизонтах Земли: в нижней атмосфере, в гидросфере, в почвах, в глубинах литосферы, пропитанных природными водами и нефтяными месторождениями.
По оценкам разных источников биосфера возникла 3,5–4,5 млрд лет назад. Область современного обитания живых организмов по вертикали охватывает в среднем 10–20 км; сфера случайного попадания организмов и биогенных элементов колеблется до 40–50 км. Считается, что нижняя граница биосферы в среднем лежит на глубине 3 км от поверхности суши и 0,5 км ниже дна океана, хотя в буровых скважинах живые микроорганизмы обнаружены на глубине 4 км, а микробиологические остатки – до 7 км. В выходах термальных вод на дне океана на глубинах в 3 км при давлении около 300 атм. (3•107 Па) обнаружены живые организмы при температуре 2500С. Растения поднимаются в горы до высоты более 5 км. Дальше царствует вечный холод, но и здесь присутствует жизнь – обитают микроорганизмы. Верхняя граница биосферы находится на высоте 20–25 км на уровне озонового слоя, выше случайно залетают только споры бактерий и грибов.
По современным вероятностным оценкам общее количество массы живого вещества в современную эпоху составляет порядка 2420 млрд т. Эту величину можно сравнить с массой оболочек Земли, в той или иной степени охваченных биосферой (табл. 2.1).
| Таблица 2.1 | ||
| Сравнение масс оболочек Земли | ||
| Оболочки Земли | Масса, т | Отношение к массе живого вещества |
| Живое вещество | 2,4 • 1012 | |
| Атмосфера | 5,15• 1015 | 2 146 |
| Гидросфера | 1,5 • 1018 | 625 000 |
| Земная кора | 2,8 • 1019 | 11 670 000 |
Таким образом, все живое вещество нашей планеты составляет примерно 1/10 000 000 часть массы земной коры. Однако в качественном отношении живое вещество представляет собой наиболее высокоорганизованную часть материи Земли. По своему активному воздействию на окружающую среду оно занимает особое место и качественно резко отличается от других оболочек земного шара, так же, как живая материя отличается от мертвой. Оценка среднего химического состава живого вещества была произведена А.П. Виноградовым. Из таблицы 2.2 видно, что главные составные части живого вещества – это элементы, широко распространенные в природе: в атмосфере, гидросфере, литосфере и космосе. Средний элементарный состав живого вещества отличается от состава земной коры высоким содержанием углерода. По содержанию других элементов организмы не повторяют состава среды своего обитания. Они избирательно поглощают элементы, необходимые для построения их тканей. В процессе жизнедеятельности организмы используют наиболее доступные атомы, способные к образованию устойчивых химических связей. Атомы углерода имеют способность создавать длинные цепи соединений с другими атомами, что приводит к построению бесчисленных полимеров и других сложных органических высокомолекулярных соединений.
| Таблица 2.2 Средний химический состав живого вещества (по A.П. Виноградову) | |||||||
| Элемент | % | Элемент | % | Элемент | % | Элемент | % |
| О | 70,0 | Cl | 2•10-2 | Zn | 5•10-4 | As | 3•10-5 |
| С | 18,0 | Fe | 1•10-2 | Rb | 5•10-4 | Со | 2•10-5 |
| Н | 10,5 | А1 | 5•10-3 | Сu | 2•10-4 | Li | 2•10-5 |
| Са | 5•10-1 | Ba | 3•10-3 | Вг | 1,5•10-4 | Mo | 1•10-5
Дата добавления: 2015-03-03; просмотров: 2318; |
Генерация страницы за: 0.117 сек.