Современная кнцепция гена
Как отмечено выше, термин «ген» впервые для обозначения единиц наследственности был введен в литературу в 1909 г. английским генетиком У. Бэтсоном. С самого начала развития хромосомной теории наследственности представления о генах сводились к тому, что последние являются не только единицами наследственности, но и физическими единицами, располагающимися в линейном порядке на хромосомах. В рамках этих представлений в 1950 г. всеобщее признание имели следующие положения:
1. Ген является крайней единицей структуры. В соответствии с этим положением признавали, что ген нельзя разделить на субъединицы разрывом хромосом или рекомбинацией.
2. Ген является крайней единицей функции (фенотипического различия). Это положение означало, что ген детерминирует одиночную специфическую функцию в метаболизме или развитии организмов, причем любая специфическая функция не может быть «разделена» на субфункции.
На основе этих положений в то время была сформулирована гипотеза «один ген — один фермент». Между тем позднее оказалось, что белки имеют сложную полипептидную природу. В частности, известны белки, полипептидные цепи которых различны. Например, фермент Е. coli триптофансинтетаза, которая катализирует накопление индола и серина, являющихся предшественниками трип-тофана, сформирован таким образом, что одна часть молекулы имеет участок (называемый А), который соединяется с индоловым компонентом индоглицерофосфат, тогда как другая часть имеет участок (называемый В), соединяющийся с сериновым компонентом (серии + + пиридоксальфосфат). Синтез частей А и В-триптофансинтетазы детерминирован различными генами.
Данные о тонком строении генов свидетельствуют, что гипотеза «один ген — один фермент» сохраняет свое содержание и остается справедливой, но лишь в случае ферментов, представляющих собой одиночный полипептид. Данные о тонком строении генов лучше всего укладываются в постулаты «один ген — РНК одного типа, одно семейство родственых полипептидов» или «один ген — одна мРНК, один полипептид». Однако этот постулат имеет исключения, связанные с особенностями фенотипического проявления некоторых генных мутаций. В частности, недавно установлено, что мутации в тиро-зинкиназном протоонкогене человека сопровождаются одновременными изменениями четырех фенотипических функций, т. е. одновременным развитием четырех болезней (семейной мозговой ти-роидной карциномы, множественной эндокринной неоплазии типа 2А, более острой множественной эндокринной неоплазии типа 2В и болезнью Гиршенранга, характеризующейся отсутствием парасимпатической иннервации толстого кишечника). Следовательно, эти данные могут укладываться в положение «один ген — несколько признаков».
Как уже отмечено, большинство эукариотических генов являются однокопийными. Тем не менее известны гены, представленные множественными копиями. Например, гены, кодирующие белки-гистоны, у организмов ряда видов существует почти в тысяче копий, что объясняется необходимостью очень быстрого синтеза гистонов в период эмбрионального развития этих организмов. В большом количестве копий существуют гены, кодирующие синтез белка кератина (основного строительного материала перьев) у кур. Наконец, у всех исследованных позвоночных отмечено наличие значительного количества генов, кодирующих синтез четырех рибосомных РНК.
Размеры (длину) генов измеряют количеством входящих в его состав пар азотистых оснований или в единицах длины. Они весьма различны, но в среднем составляют примерно 1000—1050 пар азотистых оснований или около 357 нм (если исходить из расстояния в 0,34 нм между парами оснований в двойной спирали ДНК (0,34´1050 = 357 нм). Ген возможно охарактеризовать и в плане его массы. Поскольку молекулярная масса одной нуклеотидной пары в среднем равна 650, то масса среднего по размерам гена будет составлять 650´1050 =
= 680000. Однако известны у отдельных организмов и очень малые гены, размеры которых составляют всего лишь несколько десятков пар азотистых оснований. Например, у Е. faecalis длина отдельных генов составляет всего лишь 66 пар азотистых оснований, а у S. aureus — 78 азотистых оснований. Заметим, что средние размеры генов у Е. coil составляют, примерно, 1000 - 1100 пар азотистых оснований. Таким образом, в молекулярном плане ген представляет собой сегмент ДНК (последовательность азотистых оснований) определенной длины и массы, который контролирует синтез одного полипептида.
Однако гены кодируют не только полипептиды. Отдельные гены кодируют тРНК и рРНК. Поскольку одно азотистое основание тРНК или рРНК кодируется одним азотистым основанием ДНК, то гены, кодирующие тРНК и рРНК, характеризуются значительно меньшими размерами в сравнении с генами, кодирующими полипептиды.
В научной литературе ген часто аналогизируют с цистроном, под которым понимают наименьшую единицу наследственности. Между геном и цистроном действительно нет существенной разницы, в результате чего эти термины используют на равных правах.
Новейшие представления о тонком строении генов нашли отражение в созданных с помощью молекулярно-генетических методов физических, генетических и цитологических картах геномов растений и животных ряда видов, а также человека. Достоверность представлений о размерах и молекулярной организации генов полностью подтверждена данными генной инженерии.
Вопросы для обсуждения
1. Универсальны ли законы Г. Менделя, применимы ли они к человеку?
2. Как вы понимаете альтернативные (контрастирующие) признаки, аллель и пару аллелей?
3. Г. Мендель обнаружил, что скрещивания двух чистолинейных растений садового гороха, одно из которых давало семена круглой формы, другое — шероховатые, привели к получению гибридов, дававших семена круглой формы. Скрещивания гибридов первого поколения между собой дали начало растениям, семена которых имели круглую или шероховатую форму. Соотношение между этими растениями было 3:1.0 чем свидетельствовали эти результаты?
4. В чем заключались использованные Г. Менделем анализирующие скрещивания и какова их роль?
б. Можно ли предсказать наследование признака, контролируемого двумя аллелями?
6. Что такое наследственность, сцепленная с полом? Приведите примеры наследования пола и признаков, сцепленных с полом.
7. Что такое группы сцепления и как гены располагаются на хромосомах?
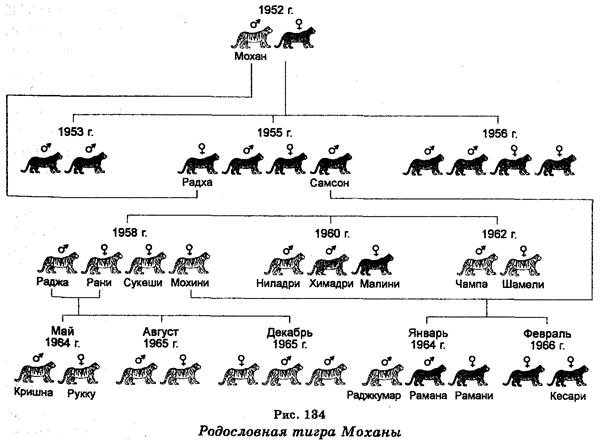
8. В 1952 г. в одном из районов Индии был пойман белый тигр (самец), которого затем скрестили с желтым тигром. Потомство от этого скрещивания приведено на рис. 134. Руководствуясь этой родословной, дайте ответы на следующие вопросы: а) Если допустить, что окраска шерсти контролируется парой аллелей, то является ли этот признак доминантным или доминантность отсутствует? б) Если окраска шерсти является примером наследования доминантности, то какая окраска (белая или желтая) является доминантным признаком? в) Определите и запишите, используя какие-либо символы, генотипы тигров Мохана, Самсона, Мохини, Малини и Кришны. г) Каково ожидаемое отношение потомства с желтым шерстным покровом к потомству с белой шерстью, полученного после скрещивания Самсона и Мохини? д) Какова вероятность рождения четырех желтых и одного белого тигрят в скрещиваниях Самсона и Мохини?
9. В семье N родились и выросли 16 детей. В табл. 18 приведены некоторые признаки, характерные для каждого из детей, когда они стали взрослыми. Используя эту таблицу, проделайте следующую работу:
а) Установите для каждого из четырех признаков тип наследования (доминантный, рецессивный, полу доминантный, отсутствие доминантности, множественность аллелей, полигенность и т. д.). б) Определите возможные генотипы и фенотипы по каждому признаку родителей N (отца и матери), в) Определите вероятность, с которой родились девочки и мальчики в этой семье.
10. Сформулируйте современную концепцию гена. Существует ли разница между геном и цистроном?
11. Что собой представляет мозаичность генов?
12. Гриб Pkytophthora castorium, который паразитирует на растениях многих видов, размножается бесполым путем с помощью подвижных зооспор, формируемых в спорангиях на мицелии, и половым путем благодаря ооспорам, которые формируются слиянием женских и мужских гамет, тоже формируемых на мицелии. Было найдено, что ооспоры, сформированные в культуре этого гриба после обработки его нитрозогуанидином, неодинаковы по своим свойствам. Одна четверть ооспор давала метионинзависимые колонии, тогда как другие три четверти ооспор напоминали родительский метиониннезависимый штамм. Анализ дальнейших генераций ооспор показал, что все метионинзависимые культуры продуцировали лишь метионинзависимые ооспоры, тогда как метиониннезависимые культуры первой генерации распадались на два класса. Около % культур характеризовались отношением около 3 : 1 метиониннезависимых и ме-тионинзависимых ооспор, тогда как отстающая треть вела себя в дальнейших скрещиваниях как фракция метиониннезависимых культур. Ооспоры в каждом случае давали начало колониям, напоминающим колонии тех культур, в которых они были сформированы независимо от того, нуждались они в метионине или нет. Каково ваше заключение о гаплоидном/ диплоидном статусе жизненного цикла грибов? В какое время развития у грибов происходит мейоз?
Таблица 18
Некоторые признаки, характерные для детей в семье
| ИМЯ | Способность ощущать вкус фенил-тиокарбамида | Группа крови по антигенам | . Группа крови по антигенам АВ, А, В, 0 | Роет (в см) |
| Иван | + | MN | АВ | |
| Наталья | - | MN | ||
| Елена | + | N | АВ | |
| Ирина | + | MN | А | |
| Валерий | + | N | В | |
| Владимир | + | М | А | |
| Андрей | - | MN | В | |
| Петр | - | N | ||
| Николай | + | MN | о | |
| Клавдия | + | MN | АВ | |
| Павел | + | М | В | |
| Людмила | + | N | А | |
| Галина | - | N | ||
| Виктор | + | MN | АВ | |
| Василий | + | MN | В | |
| Федор | + | М | А |
13. Что вы понимаете под кроссинговером и генетической рекомбинацией? Каково происхождение кроссинговера?
14. Когда две рецессивные мутации айв теплены с рекомбинант-ной фракцией генов, будет формироваться два вида двойных гетерози-гот: цис (ав)/(++) и транс (а+)/(+в). Сформулируйте предсказание относительно фенотипических отношений между потомством, полученным самоопылением или сиб-скрещиванием по каждому из этих видов двойных гетерозигот.
Литература
Гаисинович А. Е. Зарождение генетики. М.: Наука. 1997. 195 стр
Дубинин Н. П. Генетика. Кишинев: Штииица. 1985. 533 стр.
Пехов А. П. Биология и общая генетика. М.: РУДН. 1993. 439 стр
Orel V. Gregor Mendel: The First Geneticist. Oxford University Press 1995 353 pp
Дата добавления: 2015-03-03; просмотров: 878;