Тема 14. Пути ресинтеза АТФ
План:
1. Анаэробный гликолиз. Ресинтез АТФ в процессе гликолиза. Факторы, влияющие на протекание гликолиза.
2. Аэробный путь ресинтеза АТФ. Особенности регуляции.
3. Ресинтез АТФ в цикле Кребса.
4. Молочная кислота, ее роль в организме, пути ее устранения.
5. Биологическое окисление. Синтез АТФ при переносе электронов по цепи дыхательных ферментов.
1-й вопрос
Распад глюкозы возможен двумя путями. Один из них заключается в распаде шестиуглеродной молекулы глюкозы на две трехуглеродные. Этот путь называется дихотомическим распадом глюкозы. При реализации второго пути происходит потеря молекулой глюкозы одного атома углерода, что приводит к образованию пентозы; этот путь называется апотомический.
Дихотомический распад глюкозы (гликолиз) может происходить как в анаэробных, так и аэробных условиях. При распаде глюкозы в анаэробных условиях в результате процесса молочнокислого брожения образуется молочная кислота. отдельные реакции гликолиза катализируют 11 ферментов, образующих цепь, в которой продукт реакции, ускоряемой предшествующим ферментом, является субстратом для последующего. Гликолиз условно можно разбить на два этапа. В первом происходит затарта энергии, второй – характеризуется накоплением энергии в виде молекул АТФ.
Химизм процесса представлен в теме «Распад углеводов» и заканчивается переходом ПВК в молочную кислоту.
| CH3 | — | C | — | COOH | 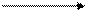
| + НАД · Н2 | CH3 | — | CН | — | COOH | |||
| || | - НАД | | | ||||||||||||
| O | ОН |
Бóльшая часть молочной кислоты, образующейся в мышце, вымывается в кровеносное русло. Изменению рН крови препятствует бикарбонатная буферная система: у спортсменов буферная емкость крови повышена по сравнению с нетренированными людьми, поэтому они могут переносить более высокое содержание молочной кислоты. Далее молочная кислота транспортируется к печени и почкам, где почти полностью перерабатывается в глюкозу и гликоген. Незначительная часть молочной кислоты вновь превращается в пировиноградную кислоту, которая в аэробных условиях окисляется до конечного продукта.
2-й вопрос
Аэробный распад глюкозы иначе называется пентозофосфатным циклом. В результате протекания этого пути из 6 молекул глюкозо-6-фосфата распадается одна. Апотомический распад глюкозы можно разделить на две фазы: окислительную и анаэробную.
Окислительную фазу где глюкозо-6-фосфат превращается в рибулёзо-5- фосфат представлена в вопросе «Распад углеводов. Аэробный распад глюкозы»
Анаэробная фаза апотомического распада глюкозы.
Дальнейший обмен рибулозо-5-фосфата протекает очень сложно, имеет место превращение фосфопентоз – пентозофосфатный цикл. В результате которого из шести молекул глюкозо-6-фосфата, вступающих в аэробный путь распада углеводов одна молекула глюкозо-6-фосфата полностью расщепляется с образованием СО2, Н2О и 36 молекул АТФ. Именно наибольший энергетический эффект распада глюкозо-6-фосфата, по сравнению с гликолизом (2 молекулы АТФ), имеет важное значение в обеспечении энергией мозга и мышц при физических нагрузках.
3-й вопрос
Цикл ди- и трикарбоновых кислот (цикл Кребса) занимает важное место в процессах обмена веществ: здесь идет обезвреживание ацетил-КоА (и ПВК) до конечных продуктов: углекислого газа и воды; синтезируется 12 молекул АТФ; образуется ряд промежуточных продуктов, которые используются для синтеза важных соединений. Например, щавелевоуксусная и кетоглутаровая кислоты могут образовать аспарагиновую и глутаминовую кислоты; ацетил-КоА служит исходным веществом для синтеза жирных кислот, холестерина, холевых кислот, гормонов. Цикл ди- и трикарбоновых кислот является следующим звеном основных видов обмена: обмена углеводов, белков, жиров. Подробно смотри в теме «Распад углеводов».
4-й вопрос
Увеличение количества молочной кислоты в саркоплазматическом пространстве мышц сопровождается изменением осмотического давления при этом вода из межклеточной среды поступает внутрь мышечных волокон, вызывая их набухание и регидность. Значительные изменения осмотического давления в мышцах могут быть причиной болевых ощущений.
Молочная кислота легко диффундирует через клеточные мембраны по градиенту концентрации в кровь, где вступает во взаимодействие с бикарбонатной системой, что приводит к выделению «неметаболического» избытка СО2:
NаНСО3 + СН3 – СН – СООН СН3 – СН – СООNа + Н2О + СО2
ОН ОН
Таким образом, увеличение кислотности, повышение СО2, служит сигналом для дыхательного центра, при выходе молочной кислоты усиливается легочная вентиляция и поставка кислорода работающей мышцы.
5-й вопрос
Биологическое окисление – это совокупность окислительных реакций, происходящих в биологических объектах (в тканях) и обеспечивающих организм энергией и метаболитами для осуществления процессов жизнедеятельности. При биологическом окислении также идет разрушение вредных продуктов обмена веществ, продуктов жизнедеятельности организма.
В развитии теории биологического окисления принимали участие ученые: 1868 г. - Шёнбайн (немецкий ученый), 1897 г. - А.Н. Бах, 1912 г. В.И. Палладин, Г.Виланд. Взгляды этих ученых положены в основу современной теории биологического окисления. Её суть.
В переносе Н2 на О2 участвуют несколько ферментных систем (дыхательная цепь ферментов), выделяют три основных компонента: дегидрогеназы (НАД, НАДФ); флавиновые (ФАД, ФМН); цитохромы (гем Fe2+). В результате образуется конечный продукт биологического окисления – H2O. В биологическом окислении участвует цепь дыхательных ферментов.
Первый акцептор Н2 – дегидрогеназа, кофермент – либо НАД (в митохондриях), либо НАДФ (в цитоплазме).

| H(H+ē) | |||||||||||
| субстрат |
| → | дегидрогеназы | → | флавопротеины | → | цитохромы | → | ½O2 | → | H2O | |
| H(H+ē) |

| H(H+ē) |
|
|
|

|
| ||||||||||||||||||||||
| субстрат | 
| → | НАД | → | ФАД | 
| b |
| c | 
| a1 | 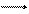 a3 a3
| ½O2 | 
| H2O | |||||||||||||
| H(H+ē) | (НАДФ) |
2ē+O → O2-
2H++O2- → H2O
Субстраты: лактат, цитрат, малат, сукцинат, глицерофосфат и другие метаболиты.
В зависимости от природы организма и окисляемого субстрата окисление в клетках может осуществляться главным образом по одному из 3-х путей.
 1.При полном наборе дыхательных ферментов, когда идет предварительное активирование О в О2-.
1.При полном наборе дыхательных ферментов, когда идет предварительное активирование О в О2-.

 H+
H+


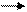 Н (Н+е-) Н+е- 2е- 2е- 2е- 2е- 2е-
Н (Н+е-) Н+е- 2е- 2е- 2е- 2е- 2е-






 S НАД ФАД b c a1 a3 1/2O2 H2O
S НАД ФАД b c a1 a3 1/2O2 H2O

 Н (Н+е-) Н+е-
Н (Н+е-) Н+е-
H+
2.Без цитохромов:
Н
 S НАД ФАД О2 Н2О2.
S НАД ФАД О2 Н2О2.
Н
3.Без НАД и без цитохромов:
Н
 S ФАД О2 Н2О2.
S ФАД О2 Н2О2.
Н
Учёные установили, что при переносе водорода на кислород при участии всех переносчиков образуется три молекулы АТФ. Восстановление формы НАД·H2 и НАДФ·H2 при переносе H2 на O2 дают 3 АТФ, а ФАД·H2 даёт 2 АТФ. При биологическом окислении образуется Н2О или Н2О2, она, в свою очередь, под действием каталазы распадается на Н2О иО2. Вода, образующаяся при биологическом окислении, расходуется на нужды клетки (реакции гидролиза) или выводится как конечный продукт из организма.
При биологическом окислении выделяется энергия, которая либо переходит в тепловую и рассеивается, либо накапливается в ~ АТФ и потом используется на все жизненные процессы.
Процесс, при котором идет накопление энергии, освободившейся при биологическом окислении, в ~ связях АТФ – окислительное фосфорилирование, то есть синтез АТФ из АДФ и Ф(н) за счет энергии окисления органических веществ:
 Е
Е
 АДФ + Ф(н) АТФ + Н2О.
АДФ + Ф(н) АТФ + Н2О.
В макроэргических связях АТФ накапливается 40% энергии биологического окисления.
Впервые на сопряжение биологического окисления с фосфорилированием АДФ указал В.А.Энгельгардт (1930 г.). Позднее В.А.Белицер и Е.Т. Цыбакова показали, что синтез АТФ из АДФ и Ф(н) идет в митохондриях при миграции е- от субстрата к О2 через цепь дыхательных ферментов. Эти ученые обнаружили, что на каждый поглощенный атом О образуется 3 молекулы АТФ, то есть в дыхательной цепи ферментов существует 3 пункта сопряжения окисления с фосфорилированием АДФ:
Н


 S НАД ФАД b c a a3 1/2О2 Н2О
S НАД ФАД b c a a3 1/2О2 Н2О




 Н
Н
АТФ АТФ АТФ
До конца механизм окислительного фосфорилирования пока не изучен. По этому вопросу выдвинут ряд гипотез: 1953 г., химическая (Липман, Слейтер); 1961 г., хемиоосмотическая (Митчел, английский ученый); 1964 г. механохимическая (конформационная) (Байер). Сейчас общепринята гипотеза Митчела, которая поддержана и получила развитие в работах В.П.Скулачева (1972 г.). Ферменты – переносчики дыхательной цепи сосредоточены в мембране (точнее – во внутренней мембране митохондрий).
Дегидрогеназы присоединяют к себе Н2 от субстратов, образующихся в результате реакций цикла Кребса (при обмене углеводов, белков, жиров). При переходе на цитохромную систему осуществляется перенос е-. При этом Н2 выбрасываются (активный перенос) из внутримитохондриального пространства (матрикса) наружу, благодаря этому создается градиент ионов водорода – градиент рН.
Н+ внешняя сторона

ОН- матрикс
Мембрана оказывается поляризованной. С наружной стороны мембраны накапливаются ионы Н+, а с внутренней – ионы ОН-. Вследствие того, что по обе стороны мембраны находятся разнозаряженные частицы возникает электрохимический мембранный потенциал, который является движущей силой для синтеза АТФ.
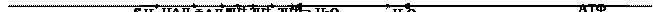
Синтез АТФ катализируется АТФ-синтетазой, расположенной в мембране.
АТФаза
 АДФ + Ф(н) АТФ + Н+ + ОН-
АДФ + Ф(н) АТФ + Н+ + ОН-
АТФ будет синтезироваться, если образующаяся вода будет удаляться. Это достигается благодаря тому, что в силу градиента рН ионы ОН- воды вытягиваются в наружнее пространство, а ионы Н+ - во внутреннее пространство митохондрий. При переносе пары е- во внешнее пространство выбрасывается 6 протонов (Н+), что приводит к образованию 3-х молекул АТФ.
Дата добавления: 2015-02-28; просмотров: 2125;