СЕНСОРНЫХ СИСТЕМ. У позвоночных животных в процессе эволюции возникло большое разнообразие сенсорных систем (анализаторов)
У позвоночных животных в процессе эволюции возникло большое разнообразие сенсорных систем (анализаторов). Благодаря этому ЦНС получает информацию о процессах в организме и событиях внешнего мира. Анализаторы различных сигналов играют неодинаковую роль у представителей разных отрядов млекопитающих. Например, приматы ориентируются во внешней среде преимущественно при помощи зрения, а обоняние играет относительно незначительную роль. У хищных ведущее значение имеют слух и обоняние и т. д. В сенсорной системе выделяют периферическую часть, представленную полем рецепторов, и центральную, образованную несколькими уровнями обработки в ЦНС, самым высоким из которых является кора больших полушарий, а также пути, которые связывают эти уровни.
Рецепторы принято делить на три больших класса: экстерорецепторы, интерорецепторы и проприорецепторы. Среди экстерорецепторов выделяют дистантные (зрительные, слуховые и пр.) и контактные (температурные, тактильные и др.). Интерорецепторы сигнализируют о состоянии внутренних органов и изменениях химического состава тканевой жидкости, содержимого пищеварительного тракта и пр. Благодаря функционированию проприорецепторов ЦНС получает информацию о состоянии опорно-двигательного аппарата (этот тип рецепторов был подробно рассмотрен выше).
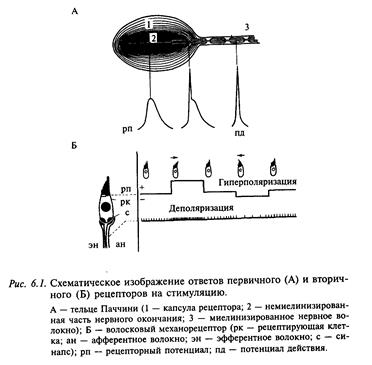
По физиологическим характеристикам рецепторы подразделяют на первично-чувствующие (или первичные) и вторично чувствующие (или вторичные). Первые преобразуют энергию внешнего стимула в возбуждение непосредственно на мембране рецептора. Примером может служить тельце Паччини (рис. 6.1, А). Рецептор представляет собой немиелинизированное окончание аксона, одетого соединительнотканной капсулой. При сдавливании рецептора от него микроэлектродом можно зарегистрировать деполяризационный потенциал, который получил название рецепторного потенциала. Деполяризация рецептора приводит к импульсному ответу в афферентном волокне. Во вторично рецепторе внешний стимул также вызывает генерацию рецепторного потенциала (рис. 6.1, Б). Афферентное волокно образует на рецепторе синапс. Рецепторный потенциал вызывает выброс медиатора из рецептора в синаптическую щель. Это приводит к генерации в афферентном волокне генераторного потенциала, а при превышении порога в афферентном волокне развивается импульсный ответ. Генераторный потенциал может быть как де-, так и гиперполяризационным и соответственно вызывать или тормозить импульсный ответ афферентного волокна. Слуховые, вестибулярные и другие типы рецепторов представлены волосковыми клетками, которые являются типичными вторичными рецепторами. Поверхность основания рецептора находится в межклеточной жидкости, и на этой поверхности локализуется синаптический контакт афферентного, а в некоторых случаях также эфферентного волокна. Благодаря последним чувствительность рецептора может управляться из нервного центра. Похожий случай был уже описан выше для мышечного веретена.
Апикальная (верхняя) поверхность волосковой клетки находится в эндолимфе с высоким содержанием калия (рис. 6.2). Строение апикальной части волоскового рецептора значительно варьирует в разных сенсорных органах, у разных видов животных, а иногда зависит от стадии развития данного вида животного. Пучок волосков на апикальном полюсе клетки состоит из 30—150 стереоцилий, которые упорядоченно располагаются на поверхности клетки. В ряде рецепторов этого типа в пучок включена также одна киноцилия, которая представляет собой измененный жгутик; она прикрепляется в цитоплазме клетки к специальной органелле — базальному тельцу.
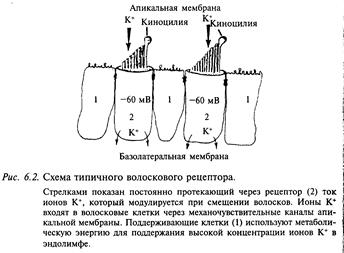
Волосковый рецептор обладает направленной чувствительностью: смещение пучка в сторону киноцилии деполяризует клетку, а смещение в противоположную сторону ее гиперполяризует (см. рис. 6.1, Б). Пороговая величина смещения волосков составляет менее 1 мкм. Ответ рецептора определяется изменением ионной проводимости апикальной мембраны. В покое клетка имеет небольшое число открытых калиевых каналов, по которым ионы калия входят в клетку по электрическому градиенту, так как внутриклеточный потенциал волосковой клетки имеет отрицательный знак относительно эндолимфы. Входящий поток катионов приводит к устойчивому входящему току, который называется рецепторным током. Этот ток деполяризует клетку примерно до уровня -60 мВ. Ионы калия, которые входят в клетку через механочувствительные калиевые каналы апикальной мембраны, покидают ее через механонечувствительные калиевые каналы основания мембраны. Через эти каналы ионы калия движутся по химическому градиенту, так как эта мембрана находится в межклеточной среде с относительно низким содержанием калия. Таким образом, через волосковый рецептор постоянно протекает рецепторный ток, величина которого модулируется смещением стереоцилий.
Все известные типы рецепторов обладают специфичностью, т. е. они наиболее эффективно возбуждаются стимулом только одной модальности. В этом смысле каждый рецептор выполняет роль фильтра, т.е. передает в ЦНС информацию только о стимулах одной модальности. Исходя из этого рецепторы можно разделить на ряд групп по модальности: механо-, термо-, хеморецепторы и др.
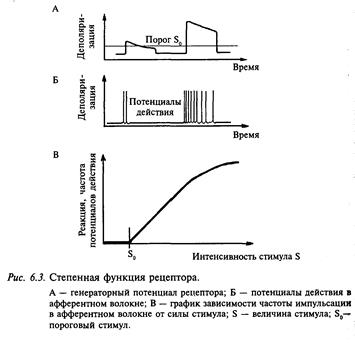
Важным понятием сенсорной физиологии является порог. Различают абсолютный и дифференциальный (разностный) пороги. Абсолютным порогом считают минимальную силу адекватного стимула, при которой возникает возбуждение рецептора. Под дифференциальным порогом понимают минимальный прирост силы стимула, который вызывает заметное изменение реакции рецептора. Внешнее воздействие кодируется (преобразуется) при помощи соответствующих рецепторов в последовательность импульсов (сенсорный код). Это преобразование в подавляющем большинстве рецепторов носит нелинейный характер. Для ряда рецепторов было установлено, что передаточная функция была показательной:
F = k(S – S0)n,
где F — частота импульсов в соответствующем афферентном волокне; k — константа; S — величина стимула; S0 — амплитуда порогового стимула; n — показатель степени (для большинства рецепторов около 1).
Это уравнение описывает широкий класс реакций рецепторов. Следует заметить, что передаточная функция рецепторов в общем случае может быть описана как «функция сжатия», так как с увеличением одной переменной (силы стимула) шкала другой (частота импульсов в афферентном волокне) все больше сжимается (рис. 6.3). Очевидно, что такая сжимающаяся шкала имеет преимущество перед линейной шкалой. «Прибор» с такой шкалой позволяет измерять величины в широком диапазоне, не слишком растягивая шкалу, за счет того что для малых величин шкала растянута, а для больших — сжата, чем и достигается удовлетворительная точность измерений как при высоких, так и при низких уровнях сигнала.
Другой широко распространенный способ кодирования в ЦНС получил название «кодирование номером линии». Этот тип кодирования хорошо прослеживается на примере передачи информации от кожи. В коже находится большое число рецепторов (рецепторы давления, температурные, болевые и др.), каждый из которых имеет собственный канал, по которому информация передается в ЦНС (подробнее см. ниже).
Большое число рецепторов спонтанно разряжаются в отсутствие каких-либо раздражителей. Например, рецепторы вестибулярного аппарата постоянно активны, что дает им возможность сигнализировать не только о величине, но и о направлении действующего стимула (например, ускорения). Для этих случаев порог определяется не как возникновение, а как изменение реакции рецептора — дифференциальный порог.
При длительном действии адекватного стимула порог срабатывания данного рецептора повышается. Это явление называется адаптацией. В адаптации рецептора принимает участие большое число процессов, и ее нельзя объяснить каким-либо одним механизмом. Например, в рецепторах кожи самым важным местом адаптации является ткань, окружающая сенсорное нервное окончание. К этой механической адаптации прибавляется адаптационный процесс генераторного потенциала. В фоторецепторах сетчатки также происходит несколько процессов: одни связаны с фотохимическими реакциями, другие — с активностью нервной сети сетчатки. В каждом рецепторе имеется свое сочетание факторов, определяющих процесс адаптации.
Отдельное афферентное нервное волокно 1-го порядка собирает информацию с более или менее широкой области поля рецепторов. Та часть рецепторной поверхности, от которой сигналы получает одно афферентное волокно, называется его рецептивным полем. Рецептивные поля соседних элементов, как правило, перекрываются. Перекрытие отдельных рецептивных полей, по-видимому, играет значительную роль. Прежде всего этим обусловливается надежность функции, которая благодаря такой организации обеспечивается большим числом рецепторов.
Одним из важных принципов взаимодействия в нейронных сетях является латеральное торможение: возбужденные элементы (рецепторы, нейроны) через возвратные коллатерали затормаживают соседние элементы. Известно, что латеральное торможение существует на всех уровнях сенсорных систем. Благодаря такому тормозному взаимодействию предотвращается «растекание» возбуждения по нервной сети, происходит своеобразное увеличение контраста, т.е. степени перепада между возбужденными и невозбужденными нейронами.
В состав сенсорной системы, кроме описанного 1-го уровня обработки информации, входит также ряд других структур головного мозга с соответствующими проводящими путями. Одними из важных станций переключения афферентной импульсации к коре служат специфические ядра таламуса. Высшим уровнем обработки сенсорных сигналов является кора больших полушарий, которая достигает наибольшего развития у млекопитающих и особенно у приматов. Кора головного мозга млекопитающих, покрывающая большие полушария конечного мозга, благодаря многочисленным складкам может иметь значительную площадь; у человека, например, она достигает 1700—2500 см В начале ХХ века была разработана классификация полей коры по признакам особенностей клеточного состава и характера миелинизации аксонов. На основании изучения клеточного состава отдельных слоев в коре выделено 11 областей, которые, в свою очередь, были разделены на 52 поля (рис. 6.4).
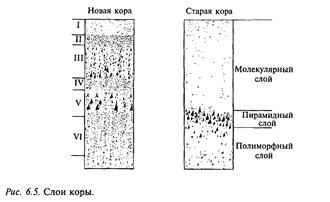
Толщина коры у млекопитающих разных видов колеблется от 1 до 6 мм. Нейроны по толщине коры распределены неравномерно и обычно образуют в новой коре 6 слоев, в старой коре — З слоя (рис. 6.5). Однако в отдельных областях новой коры количество слоев может увеличиваться или уменьшаться. Слои V и VI содержат пре имущественно крупные пирамидные нейроны, аксоны которых образуют эфферентные пути из коры. Основные афферентные пути в кору заканчиваются на нейронах верхних слоев (III и IV). Эти слои наиболее сильно развиты в центральных отделах зрительного, слухового и кожного анализаторов. По современным представлениям, такое деление слоев коры на афферентные и эфферентные нужно считать в значительной степени условным. В последние годы при изучении внутрикорковых связей установлено, что эфферентные аксоны нейронов нижних слоев образуют многочисленные возвратные коллатерали, которые восходят до самых верних слоев коры.
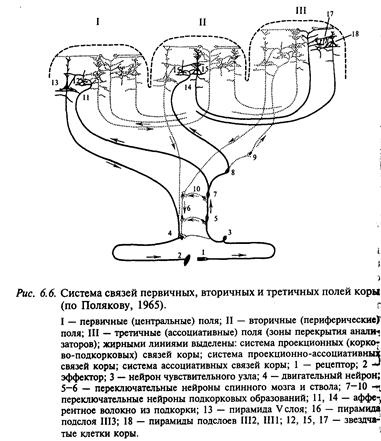
В соответствии с цитоархитектоническими и нейрофизиологическими данными выделяют проекционные и ассоциативные зоны коры. В проекционных зонах различают первичные, вторичные и третичные зоны. По И. П. Павлову, под ядром анализатора понимают зону коры, в которой после соответствующих переключений В подкорковых ядрах имеет место детальное представительство периферического отдела анализатора. Принцип деления на первичные, вторичные и третичные поля иллюстрируется рис. 6.6, на котором схематично показано, что возбуждение от соответствующих рецепторов направляется в первичные зоны по быстропроводящим путям, тогда как активация вторичных и ассоциативных зон коры происходит по полисинаптическим путям. Кроме этого, корковые поля связаны между собой многочисленными ассоциативными волокнами. В корковых проекциях сенсорных систем соблюдается принцип топии, например, в зрительной системе ретинотопия — проекция сетчатки — в поле 17 (первичная проекция), поле 18 (вторичная) и поле 19 (третичная). Этот принцип состоит в том, что, например, сетчатка образует проекцию на соответствующую область коры в некотором масштабе. При этом объем корковой проекции пропорционален плотности рецепторов в данном месте сетчатки. Благодаря этому fovea (зрительная ямка) сетчатки в корковой проекции представлена большей площадью, чем периферия сетчатки. Поражение первичных зон сенсорных систем всегда сопровождается грубыми нарушениями соответствующей сенсорной или моторной функции. В отличие от этого поражение вторичных или третичных зон вызывает расстройство аналитико-синтетической деятельности мозга, например гнозиса и праксиса, речи, целенаправленного поведения и пр. Подробнее см. главу 7 «Высшие функции нервной системы».
Вопросы
1. Принцип разделения рецепторов на первичные и вторичные.
2. Типы сенсорных порогов.
3. Виды кодирования сенсорного стимула в сенсорной системе.
4. Строение сенсорной системы.
Литература
Батуев А. С., Куликов Г. А. Введение в физиологию сенсорных систем. М.: Высшая школа, 1991.
Основы сенсорной физиологии/Под ред. Р. Шмидта. М.: Мир, 1984.
Сомьен Дж. Кодирование сенсорной информации. М.: Мир, 1976.
Шульговский В. В., Ерченков В. Г. Сравнительная физиология анализаторов. М.: Изд-во Моск. ун-та, 1989.
Дата добавления: 2015-02-10; просмотров: 1348;