Раздел II. СЕРДЕЧНО-СОСУДИСТАЯ СИСТЕМА
Спинномозговые нервы относятся к периферической (соматической) части нервной системы. Отходят от спинного мозга метамерно (посегментно) двумя корешками, отличающимися функциональной направленностью. Дорсальный (верхний) корешок, несущий на себе утолщение, спинномозговой ганглий (содержит чувствительные нейроны), немного отступя от спинного мозга, объединяется с вентральным (нижним) корешком (несёт аксоны двигательных и вегетативных (внутреностных) нейронов), образуя смешанный спинномозговой нерв и соединительную ветвь симпатической нервной системы (Рис.10).
После выхода из межпозвоночного отверстия каждый спинномозговой нерв делится на три основные ветви: дорсальную, вентральную и возвратную. Дорсальные ветви (смешанные) иннервируют дорсальную мускулатуру, позвонки, кожу соответствующих участков; вентральные ветви (смешанные) – мускулатуру и кожу нижней части тела и конечностей; возвратные (чувствительные) – оболочки мозга. И дорсальные, и вентральные ветви могут делиться на медиальные и латеральные ответвления, а, кроме того, в области отхождения от тела конечностей они формируют сплетения (плечевое и поясничное).
У бесчерепных (ланцетник) дорсальные корешки смешанные (содержат чувствительные и двигательные волокна), вентральные – только двигательные. Они иннервируют туловищную мускулатуру и повторяют её ассиметричное расположение в теле.
У круглоротых в вентральном корешке проходят только двигательные волокна, корешки не объединяются, соединительной ветви нет. Висцеральные волокна входят в состав обоих корешков, а, кроме того, у миног спинные и брюшные корешки чередуются.
У рыбот спинного мозга посегментно отходят спинномозговые нервы. Они образуются и ветвятся подобно нервам млекопитающих. Вентральные ветви передних спинномозговых нервов образуют плечевое сплетение, иннервирующее грудные плавники. Нервы подхвостовых сегментов формируют пояснично-крестцовое сплетение для иннервации брюшных плавников.
Отспинного мозга улягушкиотходит 10 пар спинномозговых нервов. Они образуются и ветвятся так же, как спинномозговые нервы млекопитающих. Плечевое сплетение образуется вентральными ветвями I – III нервов, поясничное – VII-X нервов.
У птицбольшая часть нервов плечевого сплетения иннервирует мышцы, связывающие грудную конечность с осевой частью тела, остальные – кожу и мышцы крыла. В задней части тела образуются три сплетения: нервы поясничного сплетения иннервируют мышцы тазового пояса и области бедра, нервы крестцового сплетения – практически всю тазовую конечность, нервы срамного сплетения, добавляя вегетативные нервные волокна от тазового сплетения, иннервируют половые пути (яйцевод или семяпровод) и клоаку. Нервы пояснично-крестцового сплетения прободают почки.
У млекопитающих все двигательные волокна выходят только через вентральные корешки, которые объединяются с дорсальными чувствительными, соединительная ветвь есть. Спинномозговые нервы, как и позвоночник, делятся на шейные, грудные, поясничные, крестцовые и хвостовые.
Шейные нервы (nn. cervicales) выходят через межпозвоночные отверстия в количестве 8-ми пар. Их дорсальные ветви иннервируют дорсальную мускулатуру (разгибатели головы и шеи) и кожу этой области. Вентральные ветви – вентральную мускулатуру (сгибатели головы и шеи), кожу. Из сплетения вентральных ветвей V,VI,VII шейных нервов образуется диафрагмальный нерв, идущий к диафрагме. Вентральные ветви V,VII и VIII шейных нервов входят в состав плечевого сплетения, отдающего нервы в грудную конечность.
Грудные нервы (nn.thoracales) своими дорсальными ветвями иннервируют дорсальную мускулатуру позвоночного столба, кожу холки и спины, вентральными (межрёберными – nn. intercostales) – стенку грудной клетки. I и II грудные нервы входят в состав плечевого сплетения.
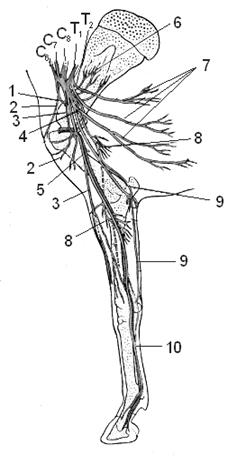
Плечевое сплетение(plexus brachialis) (Рис.11)располагается с медиальной стороны грудной конечности на уровне плечевого сустава. Парное. Образуется вышеназванными ветвями шейных и грудных спинномозговых нервов. Из него выходит 8 основных нервов:
- Надлопаточный нерв (n. suprascapularis) иннервирует разгибатели и абдукторы плечевого сустава (предостную, заостную мышцы), лопатку, плечевой сустав.
- Подлопаточный нерв (n.subscapularis) разветвляется в аддукторах и флексорах плечевого сустава (подлопаточной и большой круглой мышцах), лопатке и плечевом суставе.
- Подмышечный нерв (n. axillaris) разветвляется в области плеча и предплечья. Иннервирует сгибатели плечевого сустава (дельтовидную, большую и малую круглые мышцы), кожу латеральной поверхности плеча и предплечья.
- Мышечно-кожный нерв (n. musculocutaneus) иннервирует коракоидноплечевую и двуглавую мышцы плеча, разветвляется в коже предплечья с медиальной стороны.
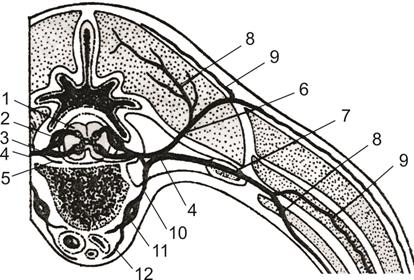
Рис.10 Ветвление спинномозгового нерва: 1 – спинной мозг,
2 – дорсальный корешок спинномозгового нерва со спинномозговым ганглием, 3 – вентральный корешок спинномозгового нерва, 4 – спинномозговой нерв, 5 – возвратная ветвь, 6 – дорсальная ветвь, 7 – вентральная ветвь, 8 – медиальная ветвь, 9 – латеральная ветвь, 10 – белая соединительная ветвь, 11 – позвоночный симпатический ганглий,
12 – симпатический нерв, 13 – тело позвонка.
- Лучевой нерв (n. radialis) – самый длинный нерв из иннервирующих разгибатели. Разветвляясь, иннервирует разгибатели локтевого (трёхглавую и локтевую мышцы, напрягатель фасции предплечья), запястного (лучевой разгибатель запястья, длинный абдуктор большого пальца) и пальцевых (общий и специальные пальцевые разгибатели) суставов, кожу предплечья и локтевой сустав. Его ветви доходят до фаланг пальцев в виде дорсальных пальцевых нервов.
- Локтевой нерв (n. ulnaris) проходит по медиальной поверхности плеча к локтевому бугру и разветвляется в мышцах запястного (локтевых сгибателе и разгибателе запястья) и пальцевых (поверхностном и глубоком пальцевых сгибателях) суставов, в плечевой и локтевой костях, коже предплечья. Конечные ветви сливаются с пальмарными нервами.
- Срединный нерв (n. medianus) – главный чувствительный нерв конечности. Проходит по медиальной поверхности плеча и предплечья в сгибатели запястья и пальцев, разветвляясь на пальмарные нервы, по ходу отдавая ветви к костям, связкам, коже.
- Грудные нервы (nn.pectorales) – делятся на краниальную группу (содержит 3-4 ветви), иннервирующую поверхностную и глубокую грудные мышцы, и каудальную группу (систоит из 4 ветвей), идущую к зубчатой вентральной и широчайшей мышце спины.
Поясничныенервы (nn. lumbales) своими дорсальными ветвями инервируют дорсальные мышцы и кожу поясницы, вентральные идут к мышцам и коже брюшной стенки, сгибателям позвоночного столба, коже мошонки и вымени, а также образуют поясничное сплетение, от которого идут нервы к тазовой конечности.
Поясничное сплетение (plexus lumbalis) (Рис.12) содержит 7 основных нервов:
|
- Подвздошно-подчревный нерв (n.iliohypogastricus) отходит от 1-2 поясничных спинномозговых нервов, идёт в большую, квадратную поясничные и брюшные мышцы, а также в кожу брюшной стенки и наружных половых органов, а у самок и в кожу вымени.
- Подвздошно-паховый нерв (n. ilioinguinalis) начинается от 2-3 поясничных нервов, иннервирует поясничные и брюшные мышцы, кожу бедра, наружные половые органы и вымя.
- Половобедренный (наружный семенной) нерв (n.genitofemoralis) отходит от 2-4 поясничных нервов, отдаёт ветви в малую, квадратную поясничные и брюшные мышцы, кожу медиальной поверхности бедра, вымя (у самок) и наружные половые органы (у самцов).
- Латеральный кожный нерв бедра (n. cutaneus femoris lateralis) отходит от 4-5 поясничного позвонка и идёт в кожу передней поверхности коленного сустава.
- Бедренный нерв (n. femoralis) разветвляется в подвздошной и четырёхглавой мышцах бедра. На середине бедра от него ответвляется ясный нерв (n. saphenus) или подкожный нерв бедра и голени, идущий по медиальной поверхности бедра, иннервируя портняжную, гребешковую и стройную мышцы, а также кожу бедра, голени и плюсны.
- Запирательный нерв (n. obturatorius) выходит из тазовой полости через запертое отверстие и разветвляется в аддукторах тазобедренного сустава (наружной запирательной, гребешковой, стройной и приводящей мышцах).
Крестцовые нервы (nn.sacrales) выходят через дорсальные и вентральные отверстия крестцовой кости. Их дорсальные ветви иннервируют кожу и мышцы крупа, а вентральные образуют крестцовое сплетение, которое соединяется с поясничным в единое пояснично-крестцовое сплетение. Из него идут нервы к тазовой конечности, наружным половым органам, мышцам ануса и хвоста.
Крестцовое сплетение (plexus sacralis) (Рис.12) отдаёт 6 основных нервов:
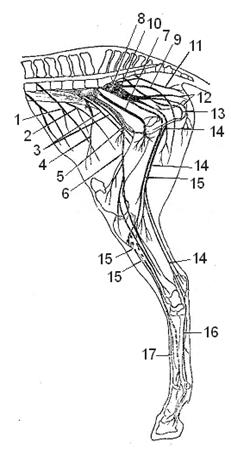
Рис.12 Пояснично-крестцовое сплетение лошади. Поясничное сплетение: 1 – подвздошноподчревный нерв, 2 – подвздошнопаховый нерв, 3 – половобедренный нерв, 4 – латеральный кожный нерв бедра, 5 – бедренный нерв, 6 – ясный нерв, 7 – запирательный нерв. Крестцовое сплетение: 8 – краниальный ягодичный нерв, 9 – каудальный ягодичный нерв, 10 – седалищный нерв, 11 – каудальный ректальный нерв, 12 – каудальный кожный нерв бедра, 13 – срамной нерв, 14 – большеберцовый нерв, 15 – малоберцовый нерв, 16 – плантарные плюсневые нервы. 17 – дорсальные плюсневые нервы.
- Краниальный и каудальный ягодичные нервы (nn. gluteus cranialis et caudalis) иннервируют ягодичные мышцы и отдают ветви в двуглавую мышцу бедра.
- Седалищный нерв (n.ischiadicus) - самый толстый и длинный нерв крестцового сплетения. Иннервирует глубокие мышцы тазобедренного сустава, проходит через большую седалищную вырезку и делится на большеберцовый и малоберцовый нервы: большеберцовый нерв (n. tibialis) иннервирует разгибатели тазобедренного (двуглавую, полусухожильную и полуперепончатую мышцы) и заплюснового (трёхглавую мышцу голени) суставов и сгибатели пальцев, а также кости, связки и кожу. Дистально он переходит в плантарные плюсневые и пальцевые нервы, достигая копытец. Малоберцовый нерв (n.fibularis, peroneus) иннервирует сгибатели заплюсневого сустава (переднюю большеберцовую и малоберцовые мышцы), разгибатели пальцев, связки, кости и кожу этой области.
- Каудальный кожный нерв бедра (n.cutaneus femoris caudalis) иннервирует мышцы заднего контура бедра – двуглавую и полусухожильную.
- Срамной нерв (n. pudendus) у самцов простирается до головки полового члена, а у самок в клитор и половые губы.
- Каудальный ректальный (геморроидальный) нерв (n. rectales caudales) идёт в прямую кишку, мышцы ануса и хвостовые мышцы.
Хвостовые нервы(n.n. caudales)насчитывают 5-6 пар.Дорсальные ветви образуют дорсальные нервы, идущие в подниматели хвоста, вентральные – в его опускатели.
Глава 3.Вегетативная нервная система
Вегетативная нервная система играет ведущую роль в поддержании постоянства внутренней среды организма и осуществляет регуляцию приспособительных реакций при охлаждении, интенсивной мышечной работе, эмоциональном напряжении, кровопотере и других неблагоприятных факторах. Она регулирует деятельность органов кровообращения, дыхания, пищеварения, выделения, размножения. Вегетативная нервная система является филогенетически более древней и проще организованной. В ней выделяют симпатический и парасимпатическийотделы. Каждый отдел устроен принципиально одинаково: состоит из центров, располагающихся в головном и/или спинном мозге, преганглионарных волокон, ганглиев и выходящих из них постганглионарных волокон. Нервы симпатической системы выходят из спинного мозга в области от 1-го грудного до 4-го поясничного сегментов. Парасимпатические нервы выходят из среднего и продолговатого мозга и из крестцового отдела спинного мозга. Симпатические и парасимпатические центры находятся под контролем гипоталамической области промежуточного мозга и коры больших полушарий. На большинство органов симпатическая и парасимпатсческая системы, будучи антагонистами, оказывают противоположное воздействие. Как привило, симпатическая система осуществляет активизирующее действие. Она обеспечивает мобилизацию, подготовку организма к усиленной деятельности. Парасимпатическая система способствует спокойному состоянию, настраивает на отдых, пищеварение, сон.
Вегетативная нервная система развита у всех позвоночных, но лучше всего изучена у млекопитающих
У ланцетника от дорсальных нервов отходят ветви к внутренностям, где есть нервные клетки и сплетения. Там происходит переключение волокон, иннервирующих именно этот орган (по принципу строения и зон иннервации эта часть нервной системы похожа на парасимпатическую). Автономная иннервация покровов и сосудов у ланцетника не обнаружена, как и нет и позвоночных ганглиев. Таким образом, симпатическая часть вегетативной нервной системы не различается.
У круглоротых строение вегетативной нервной системы слабо отличается от вегетативной нервной системы ланцетника, но появляется блуждающий нерв.
У хрящевых рыб (Рис.13) вегетативные волокна отходят от спинного мозга в каждом сегменте тела от черепа до основания хвоста с небольшим «перерывом» в передней части туловищного отдела. Переключение вегетативных волокон черепных и некоторых туловищных нервов происходит в интрамуральных ганглиях (т.е. непосредственно в органе, как свойственно парасимпатической нервной системе). Вегетативные волокна большинства туловищных нервов переключается в мелких ганглиях, расположенных рядом с позвоночником (позвоночных ганглиях), которые слабо связаны (или вовсе не связаны) друг с другом в пограничный симпатический ствол. Вегетативная иннервация кожи не обнаружена, но от позвоночных ганглиев идут волокна к сосудам мускулатуры и костей. Эти волокна не возвращаются в состав соматических нервов, так что серые соединительные ветви у хрящевых рыб не различаются. Зоны иннервации симпатической и парасимпатической нервных систем у хрящевых рыб не перекрываются.
У костистых рыб и амфибий строение вегетативной нервной системы очень сходно. Большинство спинномозговых нервов несут в себе от спинного мозга симпатические волокна, которые переключаются в позвоночных ганглиях. Постганглионарные волокна позвоночных ганглиев входят опять в спинномозговые нервы как серые соединительные ветви, которые осу-
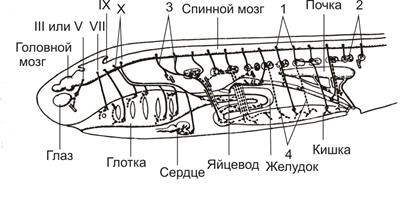
Рис.13 Вегетативная нервная система акулы
А – головной мозг, Б – спинной мозг, III - X – черепные нервы,
1 – центры, 2 – ганглии, 3 – преганглионарные волокна (_____),
4 – постганглионарные волокна (-----).
ществляют иннервацию сосудов и кожи. Позвоночные ганглии заметно соединены друг с другом в пограничный ствол, от его передней части осуществляется симпатическая иннервация головы. Переключение вегетативных волокон, отходящих от спинного мозга в составе задних туловищных нервов, происходит на нервных клетках в стенках мочевого пузыря и задней части кишечника. Таким образом, эти волокна можно отнести к парасимпатической части вегетативной нервной системы.
У костистых рыб зоны иннервации симпатической и парасимпатической систем уже частично перекрываются, далее у тетрапод количество органов с двойной иннервацией растёт.
Строение вегетативной нервной системы рептилий, птиц и млекопитающих (Рис.14) практически идентично (но у млекопитающих, например, волокна ресничного ганглия иннервируют сфинктер зрачка, а у остальных тетрапод – дилятатор).
Симпатическая нервная система. Центры симпатической нервной системы находятся в латеральных рогах серого вещества грудного и поясничного отделов спинного мозга. Преганглионарные миелиновые нервные волокна направляются от центров в составе вентрального корешка спинномозгового нерва. Вскоре после выхода через межпозвоночное отверстие вегетативные волокна отделяются от нерва в виде белой соединительной ветви и идут к ганглиям. Ганглии симпатической нервной системы по положению делятся на позвоночные и предпозвоночные
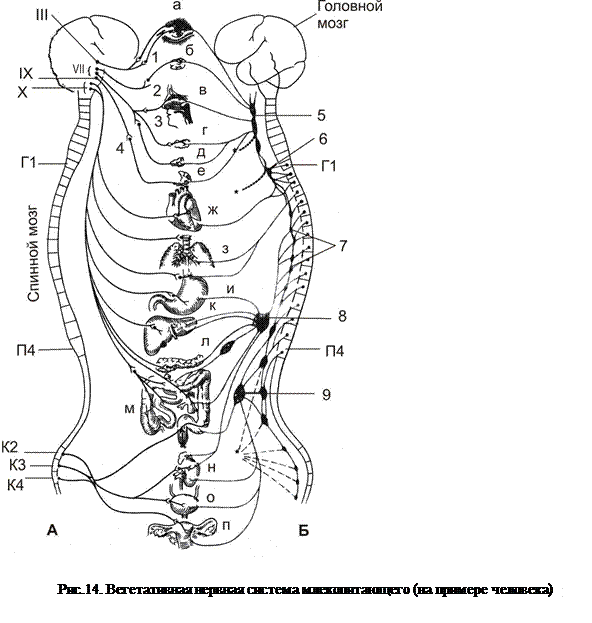
Рис.14. Вегетативная нервная система млекопитающего (на примере человека): А – парасимпатическая система, Б – симпатическая система, III – глазодвигательный нерв,VII – лицевой нерв, IX – языкоглоточный нерв, X – блуждающий нерв, Г1 – грудной нейросегмент, П4 – поясничный нейросегмент, К2 - К4 – крестцовые сегменты, 1 – ресничный ганглий, 2 – крылонёбный ганглий, 3 – подчелюстной ганглий, 4 – ушной ганглий, 5 – краниальный (верхний) шейный ганглий, 6 – каудальный (нижний) шейный ганглий, 7 – ганглий симпатического ствола, 8 – чревный ганглий и сплетение, 9 – каудальный (нижний) брызжеечный ганглий, а – глаз, б – слёзная железа, в – носовая полость, г – подчелюстная железа, д – подъязычная железа, е – околоушная железа, ж – сердце, з – лёгкие, и – желудок, к – печень, л – поджелудочная железа, м – тонкий и толстый кишечник, н – почка, о – мочевой пузырь, п – органы размножения.
. Позвоночные ганглии располагаются с двух сторон под телами позвонков. В грудном и поясничном отделах их количество соответствует числу костных сегментов. В шейном отделе три ганглия: краниальный, средний (у лошади его нет) и каудальный. Последний вместе с первым грудным ганглием образует звездчатый узел. К ганглиям подходят из центров преганглионарные волокна. Одни из них оканчиваются в ближайшем ганглии, вступая в синаптическую связь с его клетками, другие проходят через ганглии и кончаются в следующем или через несколько нервных узлов. В результате все ганглии одной стороны тела оказываются связанными друг с другом в пограничный симпатический ствол.
Постганглионарные безмиелиновые волокна, образованные нейритами клеток краниального шейного ганглия, разветвляются в голове вместе с черепномозговыми нервами. От звездчатого узла постганглионарные волокна идут к сердцу, трахее, бронхам, сосудам грудной конечности и по шее в виде позвоночного нерва, от которого отходят ветви к шейным спинномозговым нервам.
От других ганглиев постгланглионарные волокна в виде серой соединительной ветви идут к спинномозговым нервам и вместе с ними достигают иннервируемых участков тела (оболочки сосу-
дов, мышц – поднимателей волос, желез, кожи) либо отходят самостоятельно к внутренним органам.
Предпозвоночные ганглии непарные – это полулунный и каудальный брыжеечный ганглии. Полулунный ганглий образован двумя чревными и краниальным брыжеечным узлами, лежит на аорте в месте отхождения от нее чревной и краниальной брыжеечной артерий. Часть преганглионарных волокон, пройдя без изменения через ганглии пограничного симпатического ствола, достигает полулунного ганглия в виде большого и малого внутренностных нервов.
Постганглионарные волокна, в большом количестве отходящие от полулунного ганглия к желудку, кишечнику, печени, поджелудочной железе, надпочечникам, почке, селезенке, образуют солнечное (брюшное аортальное) сплетение. От каудального брыжеечного ганглия постганглионарные волокна идут к прямой кишке, органам тазовой полости и вымени, образуя тазовое сплетение.
Парасимпатическая система. Центры парасимпатической части вегетативной нервной системы находятся в ядрах среднего и продолговатого мозга, в латеральных рогах серого вещества крестцового отдела спинного мозга. Преганглионарные волокна отходят от центров в составе черепномозговых или спинномозговых нервов. Достигнув ганглиев, парасимпатические нервные волокна отъединяются от соматических нервов и входят в ганглии, расположенные вблизи или внутри иннервируемых органов. Постганглионарные волокна осуществляют парасимпатическую иннервацию.
От центров, расположенных в среднем мозге, преганглионарные волокна в составе глазодвигательного нерва доходят до ресничного узла, а от него идут постганглионарные волокна к глазу, где разветвляются в сфинктере зрачка и ресничной мышце, обеспечивая его сужение.
От центров, расположенных в продолговатом мозге, парасимпатические нервы идут четырьмя путями: 1) слезоотделительный путь начинается от ядер дна четвертого мозгового желудочка. Преганглионарные волокна в составе лицевого нерва доходят до клинонёбного ганглия, лежащего в клинонёбной ямке. Постганглионарные волокна достигают слезных желез, желез нёба и носовой полости; 2) краниальный (оральный) слюноотделительный путь начинается от ядер дна четвертого мозгового желудочка. Преганглионарные волокна в составе лицевого нерва доходят до подъязычного (подчелюстного) узла, расположенного около слюнных желез. Постганглионарные волокна входят в подъязычнуюи подчелюстнуюслюнные железы; 3) каудальный (второй) слюноотделительный путь начинается от ядер дна четвертого мозгового желудочка. Преганглионарные волокна в составе языкоглоточного нерва доходят до ушного ганглия. Постганглионарные волокна идут к околоушной слюнной железе, щечным и губным железам; 4) висцеральный путь начинается от ядер продолговатого мозга, формирующих блуждающий нерв (n. vagus). Основная масса волокон, образующих вагус, – парасимпатические волокна. Из черепной полости вагус выходит через рваное отверстие. В области шеи он идет вместе с шейным участком пограничного симпатического ствола, формируя вагосимпатикус (п. vagosimpaticus). При входе в грудную полость блуждающий нерв отделяется от симпатического и отдает соматические ветви в виде возвратного нерва к глотке и гортани. Парасимпатические ветви вагуса вместе с симпатическими образуют сплетения во всех органах грудной полости.
Вагус, сопровождая пищевод двумя стволами (дорсальный и вентральный), входит в брюшную полость и образует сплетения вместе с симпатическими нервами солнечного сплетения. Парасимпатические ганглии и постганглионарные волокна находятся в стенках иннервируемых органах (интрамурально).
Из крестцового центра преганглионарные волокна выходят со спинномозговыми крестцовыми нервами. Выйдя из позвоночного канала, они отделяются от соматических нервов и образуют тазовые нервы. Эти нервы идут к ободочной и прямой кишкам, мочевому пузырю, половым органам и достигают ганглиев, расположенных в стенках данных органов. Постганглионарные волокна осуществляют их парасимпатическую иннервацию.
Контрольные вопросы
к Разделу I «Нервная система»
1. Принципиальное строение головного мозга, деление его на отделы.
2. Изменение строения заднего отдела головного мозга у разных классов типа хордовые.
3. Изменение строения среднего отдела головного мозга у разных классов типа хордовые.
4. Изменение строения промежуточного отдела головного мозга у разных классов типа хордовые.
5. Изменение строения конечного отдела головного мозга у разных классов типа хордовые.
6. Строение спинного мозга.
7. Принципы образования и ветвления спинномозгового нерва.
8. Образование и состав плечевого сплетения.
9. Образование и состав пояснично-крестцового сплетения.
10. Принципы строения и функционирования вегетативной нервной системы.
Раздел II. СЕРДЕЧНО-СОСУДИСТАЯ СИСТЕМА
Все хордовые животные имеют замкнутую сосудистую систему, то есть кровь течёт по сосудам, нигде не изливаясь в ткани или в полости тела.
Кровь является важнейшей транспортной системой организма, разнося по всему телу животного кислород, питательные и биологически активные вещества, доставляя продукты распада к местам выведения их из организма. Очевидно, для выполнения этой функции кровь должна циркулировать – двигаться по организму в определённом и постоянном направлении. Это обеспечивается пульсацией более или менее сложной мышечной структуры, лежащей по ходу сосудов – сердца(лат. cor, cardia). При этом сосуды, приносящие кровь к сердцу, получают название «вены», а выносящие кровь из сердца – «артерии».
Глава 4. Строение сердца и круги кровообращения первичноводных хордовых
Ланцетник не имеет единого сердца, функцию насосов выполняют пульсирующие участки крупных сосудов. Наиболее развиты такие «артериальные сердца» в приносящих жаберных артериях, они создают давление, необходимое для наполнения кровью мелких жаберных капилляров, где происходит насыщение крови кислородом (газообмен у ланцетника происходит также и в покровах тела). Схема кровеносной системы, тем не менее, соответствует общему плану для первичноводных позвоночных. Венозная кровь с передней половины тела собирается в парные краниальные кардинальные вены, с задней (кроме хвоста и кишки) – в каудальные кардинальные вены. В зажаберной области кардинальные вены слева и справа сливаются в кювьеровы протоки. Хвостовая вена, переходящая в подкишечную, входит в печёночный вырост, формируя воротную систему печени (здесь кровь очищается от токсинов, отдаёт часть запасаемых веществ) и продолжается от печени как печёночная вена. Кювьеровы протоки и печёночная вена сливаются, венозная кровь поступает в брюшную аорты, идущую под жаберными щелями в краниальном направлении под воздействием присасывающей силы «артериальных сердец» на жаберных артериях. От брюшной аорты отходят приносящие жаберные артерии, переходящие в выносящие жаберные артерии. Последние собираются слева и справа над жаберными щелями в корни аорты, от которых вперёд идут сонные артерии. Корни аорты сливаются в спинную аорту, разносящую артериальную кровь по всей задней половине тела.
Сердце. Оболочки сердца. В онтогенезе сердце формируется, как результат слияния и утолщения кровеносных сосудов в области шеи зародыша. В составе стенки сердца различают три слоя: внутренний - эндокард, представленный эндотелием и рыхлой соединительной тканью. Средний, собственно функциональный и наиболее толстый слой – миокард, состоящий из сердечной поперечнополосатой мышечной ткани. Наружный слой – эпикард, включает соединительную ткань и мезотелий. Эпикард является висцеральным листком серозной околосердечной оболочки – перикарда. Наличие в полости небольшого количества серозной жидкости устраняет трение, которое могло бы возникать при движениях сердца. К париетальному листку перикарда прирастает листок внутригрудной фасции и, у млекопитающих, сердечная плевра - весь этот комплекс называется «сердечная сумка»),
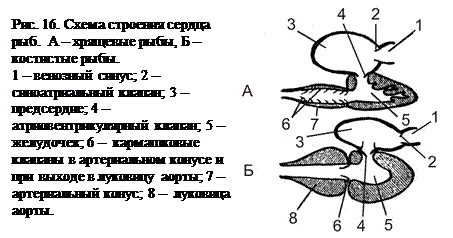 Строение сердца и круги кровообращения круглоротых и рыб. Сердце лежит каудовентрально от жаберного аппарата. Оно состоит из четырёх расположенных изогнутой цепочкой камер. Венозная (бедная кислородом) кровь по крупным венам (Кювьеровым протокам) притекает в тонкостенный венозный синус (sinus venosus). Из него кровь попадает в первую из основных камер сердца – предсердие (atrium). Оно соединено предсердно-желудочковым (атриовентрикулярным) отверстием с желудочком сердца (ventriculus). В атриовентрикулярном отверстии имеется одноимённый клапан, препятствующий обратному току крови. Желудочек с его мощным миокардом (мышечной оболочкой сердца) является основным движителем крови. Из желудочка кровь под большим давлением попадает у хрящевых рыб в артериальный конус (conus arterialis), а у круглоротых и костных рыб – в луковицу аорты (bulbus aortae), которые должны сделать ток крови по сосудам более равномерным, растягиваясь во времени силу резкого сокращения желудочка. На выходе из желудочка имеются кармашковые (полулунные) клапаны. Стенка артериального конуса имеет в своём составе миокард, сокращающийся в соответствии с общим сердечным ритмом, и несколько клапанов. Луковица аорты, не являясь частью сердца, лишена миокарда, но содержит мощный эластический слой, растягивающийся под напором крови, а затем автоматически сжимающийся, проталкивая кровь дальше, в брюшную аорту (aorta ventralis).
Строение сердца и круги кровообращения круглоротых и рыб. Сердце лежит каудовентрально от жаберного аппарата. Оно состоит из четырёх расположенных изогнутой цепочкой камер. Венозная (бедная кислородом) кровь по крупным венам (Кювьеровым протокам) притекает в тонкостенный венозный синус (sinus venosus). Из него кровь попадает в первую из основных камер сердца – предсердие (atrium). Оно соединено предсердно-желудочковым (атриовентрикулярным) отверстием с желудочком сердца (ventriculus). В атриовентрикулярном отверстии имеется одноимённый клапан, препятствующий обратному току крови. Желудочек с его мощным миокардом (мышечной оболочкой сердца) является основным движителем крови. Из желудочка кровь под большим давлением попадает у хрящевых рыб в артериальный конус (conus arterialis), а у круглоротых и костных рыб – в луковицу аорты (bulbus aortae), которые должны сделать ток крови по сосудам более равномерным, растягиваясь во времени силу резкого сокращения желудочка. На выходе из желудочка имеются кармашковые (полулунные) клапаны. Стенка артериального конуса имеет в своём составе миокард, сокращающийся в соответствии с общим сердечным ритмом, и несколько клапанов. Луковица аорты, не являясь частью сердца, лишена миокарда, но содержит мощный эластический слой, растягивающийся под напором крови, а затем автоматически сжимающийся, проталкивая кровь дальше, в брюшную аорту (aorta ventralis).
Сердце круглоротых и большинства рыб содержит только бедную кислородом венозную кровь, что соответствует наличию одного круга кровообращения. Исключениями являются двоякодышащие рыбы, о которых будет сказано позднее.
У миног и миксин сердце изогнуто во фронтальной плоскости. В венозный синус впадают парные краниальные и каудальные кардинальные вены, собирающие бедную кислородом кровь со всего организма. От желудочка отходит луковица аорты.
У рыб сердце изогнуто в сагиттальной плоскости, так, что предсердие лежит над желудочком. Вероятно, это даёт желудочку определённую «свободу хода», позволяющую ему слегка смещаться при сокращении (по Ф.Я.Дзержинскому). В венозный синус, расположенный каудодорсально от предсердия, впадают Кювьеровы протоки, образованные слиянием краниальных и каудальных кардинальных вен, и печёночная вена. Из желудочка через артериальный конус (у хрящевых рыб) и через луковицу аорты (у костных рыб) выходит брюшная аорта.
У двоякодышащих рыб, в связи с развитием лёгочного дыхания и, соответственно, появлением второго круга кровообращения, возникает необходимость в разделении потоков артериальной и венозной крови в сердце. В предсердии появляется перегородка, разделяющая его на две части: правую, в которую по кардинальным венам поступает венозная кровь, и левую, в которую лёгочные вены приносят кровь, обогащённую кислородом.
Перегородка предсердия слегка заходит в желудочек, так, что полного смешивания крови и там не происходит. В артериальном конусе клапаны формируют единую складку – спиральный клапан. Он разворачивает потоки крови так, что артериальная кровь из левой половины желудочка поступает в нижнюю часть конуса, от которой отходят две передние пары жаберных дуг, несущие кровь к голове (по сонным артериям). У некоторых двоякодышащих рыб эти артерии прерываются жабрами, где кровь дополнительно обогащается кислородом, у других жабры здесь отсутствуют. Венозная кровь из правой половины желудочка направляется в дорсальную часть конуса, а оттуда – по брюшной аорте в приносящие жаберные артерии.
Основные сосуды кругов кровообращения круглоротых и рыб.От брюшной аорты отходят парные приносящие жаберные артерии. В жабрах они расходятся на капилляры, в которых происходит газообмен между кровью и омывающей жабры водой. Выносящие жаберные артерии выше жабр объединяются в парные сосуды - корни аорты (radix aortae). К полужабре брызгальца кровь может подаваться от выносящей жаберной артерии первой дуги, то есть уже окисленная. Передняя часть корней аорты несёт кровь к головному мозгу и получает название сонных артерий (a. carotis). В них же вливается выносящая артерия первой жаберной дуги, получившая дополнительный кислород кровь от брызгальца. Каудальнее жаберного аппарата корни аорты сливаются, образуя спинную аорту (aorta dorsalis). Аорта разносит артериальную кровь по всему организму, отдавая ряд парных и непарных артерий. Из парных следует отметить артерии: подключичные артерии (a.subclavia), кровоснабжающие грудные плавники, сегментарные межрёберные артерии, васкуляризирующие позвонки, рёбра и мускулатуру, артерии, идущие к парным органам: почкам, половым железам, подвздошные артерии, направляющиеся к брюшным плавникам.
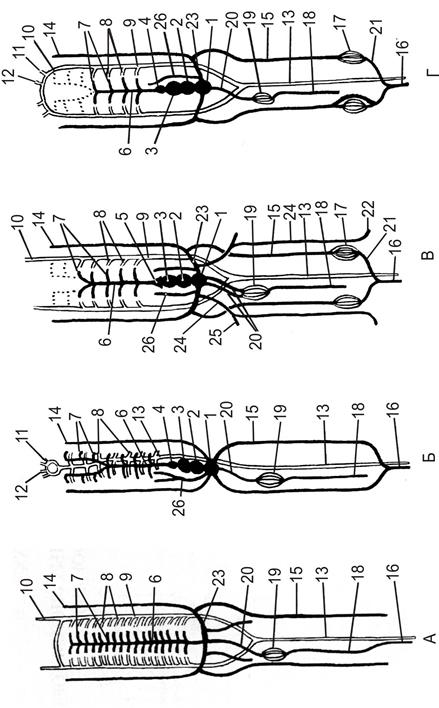 Рис. 17. Схема системы кровообращения водных хордовых. А – ланцетник; Б – круглоротые; В – хрящевые рыбы; Г – костистые рыбы.
Рис. 17. Схема системы кровообращения водных хордовых. А – ланцетник; Б – круглоротые; В – хрящевые рыбы; Г – костистые рыбы.
1 – венозный синус; 2 – предсердие; 3 – желудочек; 4 – луковица аорты; 5 – артериальный конус; 6 – брюшная аорта; 7 – приносящие жаберные артерии; 8 – выносящие жаберные артерии; 9 – корни спинной аорты; 10 – общая сонная артерия; 11 – внутренняя сонная артерия; 12 – наружная сонная артерия; 13 – спинная аорта; 14 – краниальная кардинальная вена; 15 – каудальная кардинальная вена; 16 – хвостовая вена; 17 – воротная система почек; 18 – подкишечная вена; 19 – воротная система печени; 20 – печёночная вена; 21 – воротная вена почек; 22 – подвздошная вена; 23 – Кювьеровы протоки; 24 – боковая вена; 25 – подключичная вена; 26 – ярёмная вена.
Непарные артерии идут к непарным органам, в основном системы пищеварения: желудку, кишечнику, печени, поджелудочной железе.
Каудально аорта переходит в крупную хвостовую артерию, которая проходит в канале гемальных дуг позвонков.
Венозная кровь от органов собирается в вены. От головы кровь собирается в краниальные кардинальные вены (vv. cardinales craniales), которые впадают в предсердие или самостоятельно, или предварительно объединившись с каудальными кардинальными венами в Кювьеровы протоки (ductus Cuviery). Вентрально по голове идут ярёмные вены (парные у хрящевых рыб и непарные у костистых рыб и круглоротых, v. jugularis), впадающие в предсердие самостоятельно.
По боковой поверхности тела хрящевых рыб идут парные боковые вены, впадающие в кювьеровы протоки (у круглоротых и костных рыб боковые вены не развиты).
Хвостовая вена собирает кровь от хвоста. У рыб при входе в полость тела она разделяется на две ветви - приносящие почечные артерии. В почках вены образуют воротную систему почек – разветвляются на капилляры, оплетающие канальцы нефронов. У костистых рыб одна из приносящих почечных артерий не впадает в почку или отдаёт туда небольшую по объёму ветвь. От почек отходят уже каудальные кардинальные вены (vv. cardinales caudales), идущие к сердцу.
У круглоротых воротной системы почек нет, каудальные кардинальные вены образуются из расхождения непарной хвостовой вены.
Кровь от органов пищеварения кровь и селезёнки у всех хордовых животных собирается в воротную вену печени (v.portae hepatis), которая, распадаясь в печени на капилляры, образует чудесную сеть печени. В печени кровь фильтруется, освобождаясь, в первую очередь, от аммиака. Очищенная кровь выносится печёночной веной (иногда несколькими, v. hepatice) и поступает непосредственно в предсердие.
Глава 5. Строение сердца и круги кровообращения амфибий.
Сердце амфибий трёхкамерное. Предсердия полностью разделены перегородкой. В левое предсердие впадают лёгочные вены (vv. pulmonales), несущие от лёгких богатую кислородом кровь. В венозный синус правого предсердия впадают парные краниальные полые вены (v.cava cranialis) и одна каудальная полая вена (v.cava caudalis). Каудальная полая вена несёт венозную кровь от задней части тела, краниальные полые вены образуются слиянием несущих венозную кровь яремных вен с кожными венами (v. cutanea), кровь в которых окислена (кожа амфибий активно выполняет дыхательную функцию). Таким образом, в краниальных полых венах, в венозном синусе и в правом предсердии у амфибий кровь смешанная (слабоокисленная), а не венозная.
Отверстие предсердий общее, хотя и разделённое продолжением перегородки предсердий на две половины (правую и левую). На внутренней поверхности стенок желудочка имеются высокие складки, препятствующие полному смешиванию крови из двух предсердий. Таким образом, в левой части желудочка оказывается, преимущественно, артериальная кровь, в правой — смешанная с преобладанием венозной, а в середине — смешанная кровь.
От желудочка отходит артериальный конус, который, в свою очередь, отдаёт три пары артерий: кожно-лёгочные, дуги
аорты (arcus aortae) и сонные артерии. Отверстие артериального конуса находится в правой части желудочка, поэтому при систоле (сокращении) желудочка туда последовательно поступает венозная, смешанная и, в момент максимума систолы, артериальная кровь. Разделение потоков более и менее окисленной крови по разным сосудам обеспечивается разницей в толщине и упругости стенок сосудов разных пар и работой находящегося в артериальном конусе спирального клапана (он разворачивает поток крови). Наиболее тонкие стенки и широкий просвет имеют кожно-лёгочные артерии, они и получают первую порцию крови — бедную кислородом кровь из правой части желудочка, поступающую в артериальный конус под малым давлением начала систолы. По мере усиления сокращения желудочка давление в артериальном конусе растёт и следующая порция крови — смешанная кровь центральной части желудочка — наполняет дуги аорты. Наконец при максимуме систолы открываются сонные артерии, несущие кровь к голове из левой, артериальной части желудочка.
Кожа амфибий, как орган газообмена наряду с лёгкими, образует неполный «третий» круг кровообращения. Кожная артерия, ответвляющаяся от кожно-лёгочной, несёт венозную кровь, которая в коже окисляется. Возвращается эта кровь, в отличие от артериальной крови из лёгких, не в левое, а в правое предсердие.
Особенно важное значение имеет кожная артерия и кожное дыхание для амфибий в период спячки, когда газообмен происходит практически только в покровах..

Основные сосуды кругов кровообращения амфибий.Сердечно-сосудистая система хвостатых амфибий имеет много общих черт, присущих рыбам. У взрослых бесхвостых амфибий (так называемых «высших амфибий») –– схема сосудистой системы более близка к таковой у амниот. От рыб её отличает отсутствие брюшной аорты с венозной кровью и жаберных артерий. Важной чертой является утрата каудальных кардинальных вен и формирование каудальной полой вены. Сосуды, формирующиеся при слиянии ярёмных и подключичных вен, приобретают название краниальных полых вен.
 Глава 6. Строение сердца и круги кровообращения рептилий
Глава 6. Строение сердца и круги кровообращения рептилий
Сердце рептилий имеет два полностью разделённых предсердия и один желудочек. Венозный синус и артериальный конус не развиты (за исключением гаттерии, у которой имеется венозный синус). В правое предсердие впадают две краниальные и одна каудальная полая вены, несущие венозную кровь, а в левое предсердие – лёгочные вены с артериальной кровью.
Желудочек имеет особенности строения, отличающие сердце рептилий (за исключением крокодила) от сердец других амниот. В желудочке имеется неполная перегородка, расположенная во фронтальной плоскости, таким образом, желудочек разделён на две камеры – дорсальную и вентральную. Перегородка справа не доходит до стенки желудочка, там образуется отверстие, соединяющее две половины желудочка.
Атриовентрикулярные отверстия открываются рядом друг с другом в левой половине дорсальной камеры желудочка, соответственно, левое – левее, а правое – правее. Отверстия дуг аорты располагаются практически на уровне свободного края перегородки, причём отверстие правой дуги аорты – в дорсальной камере, левой дуги – в вентральной камере. В центральной части вентральной камеры желудочка находится отверстие ствола лёгочных артерий.
Такое расположение артериальных отверстий в частично разделённом желудочке обеспечивает разделение крови на фракции с разным содержание кислорода и направление её к разным органам. При систоле предсердий кровь поступает в дорсальную камеру желудочка, при этом её левую часть – из-за расположения отверстий предсердий – заполняет артериальная кровь из левого предсердия, в правой части желудочка (у свободного края перегородки) оказывается смешанная кровь, а вентральная камера заполняется венозной кровью.
При систоле желудочка венозная кровь из вентральной камеры поступает в лёгочные артерии и направляется к лёгким для насыщения кислородом. Смешанная кровь попадает в левую дугу аорты, а правая дуга несёт практически чисто артериальную кровь. Пройдя над сердцем, дуги аорты соединяются в дорсальную аорту со смешанной кровью. Обеспечиваемое строением сердца разделение крови в дугах аорты имеет глубокий смысл, так как обе общие сонные артерии, обеспечивающие кислородом головной мозг, отходят от правой (артериальной) дуги аорты, как и обе подключичные артерии.
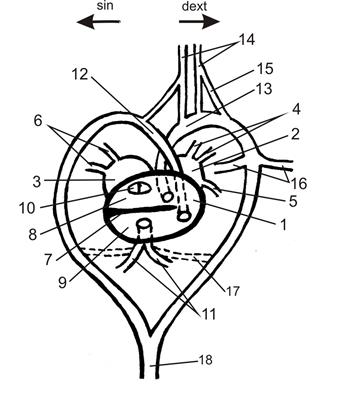
Рис. 20. Строение сердца рептилии
1 – желудочек (в сегментальном разрезе); 2 – правое предсердие; 3 – левое предсердие; 4 – краниальные полые вены; 5 – каудальная полая вена; 6 – лёгочные вены; 7 - перегородка желудочка; 8 – дорсальная камера желудочка; 9 – вентральная камера желудочка; 10 – отверстия предсердий; 11 – лёгочные артерии; 12 – левая дуга аорты; 13 – правая дуга аорты; 14 – сонные артерии; 15 – сонный проток; 16 – подключичные артерии; 17 – Боталлов проток; 18 – спинная аорта.
У рептилий сохраняются сонные протоки, соединяющие сонные артерии с соответствующими дугами аорты. Они могут быть открытыми или частично или полностью превращаться в связки.
У морских черепах сохраняется Боталлов проток, свойственный эмбриональному кровообращению всех амниот. Боталлов проток соединяет лёгочные артерии с дугами аорты, обеспечивая при нырянии переброску крови от лёгких (которые в таких условиях оказываются не поставщиками, а потребителями кислорода) в большой круг. В этом случае, единственным источником кислорода оказывается миоглобин мышц, и кровь в полых венах более оксигенирована, чем в лёгочных. Малый круг кровообращения частично запустевает, объём крови в левом предсердии оказывается небольшим и кровь из правого предсердия попадает в обе дуги аорты.
Сердце крокодила в большинстве черт соответствует строению сердца птиц и млекопитающих. В правое предсердие впадают две краниальные и одна каудальная полая вены, несущие венозную кровь, а в левое предсердие – лёгочные вены с артериальной кровью. Полная перегородка разделяет правый и левый желудочки. Желудочки практически одинаковы по размеру, одинакова и толщина их миокарда. Каждый желудочек соединяется атриовентрикулярным отверстием с соответствующим предсердием. Таким образом, в правом желудочке оказывается венозная кровь, а в левом – артериальная. От левого желудочка отходит правая дуга аорты (артериальная) и отдаёт сонные и подключичные артерии. Венозная кровь из правого желудочка поступает в малый круг кровообращения по лёгочным ар 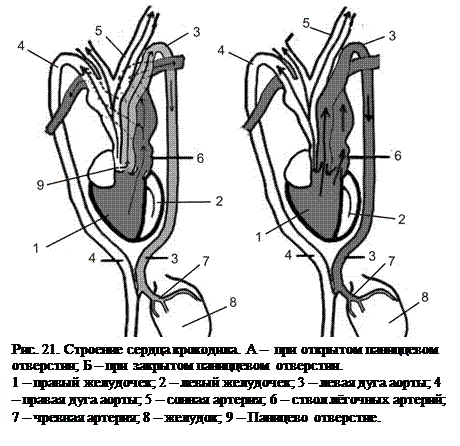 териям и в большой круг по левой (венозной) дуге аорты. Обе дуги аорты дорсокаудально от сердца соединяются в аорту, несущую смешанную кровь.
териям и в большой круг по левой (венозной) дуге аорты. Обе дуги аорты дорсокаудально от сердца соединяются в аорту, несущую смешанную кровь.
Дуги аорты в месте перекрёста – при выходе их из сердца – сообщаются с помощью Паниццева отверстия. Длительное время считали, что это отверстие служит только для переброски крови из правого желудочка в правую дугу аорты при погружении под воду, чтобы обеспечить запустевание сосудов лёгких и использовать частично оксигенированную (за счёт миоглобина мышц) кровь правого желудочка. Такой же механизм наблюдается у водных черепах, но с помощью Боталлова протока. Последними исследованиями был выявлен другой механизм, связанный с работой Паниццева отверстия. Было показано, что большую часть времени клапан Паниццева отверстия располагается так, что в обе дуги аорты поступает артериальная кровь из левого желудочка. Закрывается клапан, изолируя правую дугу и обеспечивая наличие в ней только венозной крови тогда, когда в желудок крокодила поступает крупная добыча. Углекислый газ венозной крови в идущей к желудку чревной артерии (у крокодила она отходит от левой дуги аорты, до слияния дуг) способствует резкому увеличению выработки кислоты желудочного сока, ускоряющей переваривание пищи. Вероятно, функционируют оба приведённых механизма.
Клапанный аппарат сердца крокодила соответствует клапанному аппарату сердца птиц. В правом атриовентрикулярном отверстии находится створчатый клапан, состоящий из плоских соединительнотканных створок. Створки одним краем прикреплены к краю отверстия, от свободного края отходят сухожильные струны, прикрепляющиеся к стенкам желудочка. Правое атриовентрикулярное отверстие закрывается мышечным клапаном, в артериальных отверстиях находятся кармашковые клапаны.
Дата добавления: 2015-02-10; просмотров: 1573;