Опухолеродные РНК-содержащие вирусы
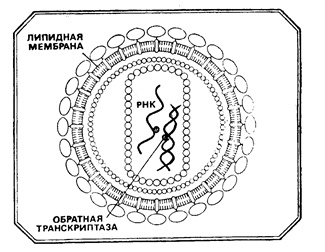
· Онкорна (RNA) вирусы.Все они являются членами общей группы ретровирусов, содержат в качестве генетического материала одноцепочечную РНК и РНК - зависимую ДНК - полимеразу (обратную транскриптазу, ревертазу).
· Обратная транскриптаза необходима для встраивания (интеграции) вирусного генома в виде ДНК - провируса в геном клетки хозяина, что обеспечивает опухолевую трансформацию. Она не требуется для поддержания опухолевого роста.
· ДНК - провирус – двунитевая ДНК, синтезированная при участии обратной транскриптазы (и других вирусных ферментов) в клеточном ядре на матрице РНК - вируса.
· Ретровирусы индуцируют опухоли и трансформируют клетки с помощью трех различных механизмов.
· Большинство ретровирусов индуцируют онкогенез через действие онкогенов. Эти вирусы могут быть четко разделены на две группы. А. Ретровирусы, несущие онкоген в составе генома, которые могут быть охарактеризованы как трансдуцирующие ретровирусы. В. Ретровирусы, не содержащие онкогены, но индуцирующие трансформацию путем интеграции в соседстве с клеточным онкогеном и последующей его активации. Такие вирусы можно охарактеризовать как цис-действующие, а саму активацию как цис-активацию (инсерционная, вставочная активация).
· Практически все трансдуцирующие ретровирусы утратили часть вирусосодержащей информации в обмен на клеточные последовательности. Поэтому они дефектны по способности синтезировать потомство, а для их репродукции необходим родственный вирус-помощник, который при совместной инфекции обеспечивает синтез необходимых компонентов вирусной частицы (белков оболочки вириона). Такие вирусы являются эффективным канцерогеном, способным трансформировать клетки в культуре и вызывать опухоли с коротким латентным периодом (несколько дней).
· Все цис-активирующие ретровирусынесодержат онкогенов, но содержат полный набор вирусных генов и способны к эффективной репликации.Они могут индуцировать опухоли (in vivo), но со значительным латентным периодом (недели, месяцы). Не способны индуцировать онкогенную трансформацию клеток in vitro. Эти вирусы не используют собственную генетическую информацию в процессе онкогенеза (в отличие от трансдуцирующих вирусов), а обеспечивают последний за счет активации клеточных генов (онкогенов), контролирующих пролиферацию клетки. К этой группе относятся вирусы ALV (лимфолейкоза птиц), ММТV (вирус опухолей молочных желез) и др.
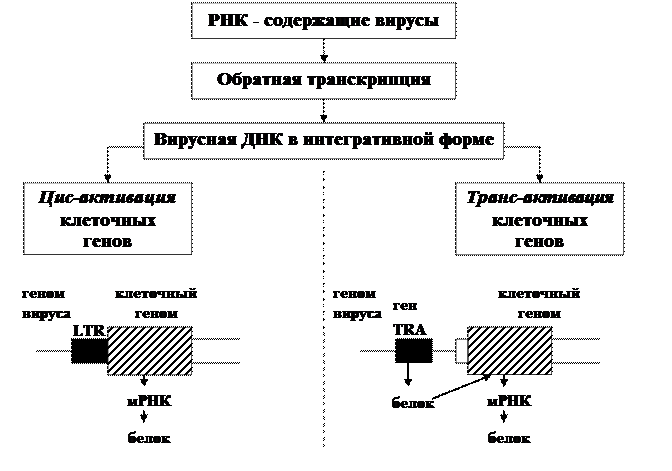
 Общий принцип взаимодействия РНК - содержащих вирусов с клетками
Общий принцип взаимодействия РНК - содержащих вирусов с клетками
(Ф.Л. Киселев, 2004)
С. Третья группа вирусов, реализующих пока еще гипотетический механизм ретровирусного канцерогенеза, имеет вирусные последовательности, которые используются не только для репликации вирусов, но и непосредственно вовлечены в канцерогенез.Эти последовательности кодируют неструктурные (т.е. не входящие в состав вирусной частицы) регуляторные белки. Данные белки могут активировать как собственные энхансерные и промоторные последовательности внутри регуляторной области вирусного генома (LTR), так и регуляторные последовательности, контролирующие активность клеточных генов, регулирующих пролиферацию. Тем самым обеспечивается возможность бесконечного деления клеток и формирования опухоли. Этот феномен получил название трансактивации.
· Трансактивация присуща не только РНК, но и ДНК-содержащим опухолеродным вирусам. Ярким примером является РНК-содержащий вирус Т-клеточного лейкоза человека (HТLV) и ДНК-содержащий вирус гепатита В (HBV).
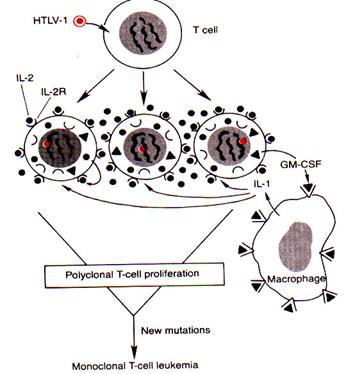 |
ПАТОГЕНЕЗ Т-КЛЕТОЧНОЙ ЛИМФОМЫ, ИНДУЦИРОВАННОЙ HTLV-1
Механизм индукции Т-клеточной лимфомы вирусом НТLV-1
(по V. Cumar, R. S. Cotran, S.L. Robbins, 1997)
· HTLV-1 не содержит v-onc и не способен к интеграции с протоонкогеном. Предполагается, что его трансформирующая активность связана с участком генома – tax. Продукт этого гена – белок taxактивирует транскрипцию клеточных онкогенов (c -fos, c – sis), генов, кодирующих ИЛ - 2, рецепторы к ИЛ - 2, а также гранулоцитарно-макрофагальный колониестимулирующий фактор – GM - CSF.
· Поскольку HTLV-1 экспрессируют ИЛ-2 и рецепторы к нему вне зависимости от субклоновой принадлежности Т-клеток, последние постоянно стимулируются к пролиферации аутокринным и паракринным путем. Происходит пролиферация всех клонов Т-лимфоцитов, т.н. поликлональная пролиферация.
· Одновременно под влиянием вируса HTLV-1 в Т-лимфоцитах происходит активация генов, контролирующих продукцию гранулоцитарно-макрофагального колониестимулирующего фактора (GM-CSF). Под действием последнего усиливается наработка клеток моноцитарно-макрофагального ряда и их активация.
· Активированные таким путем макрофаги продуцируют ИЛ-1, который оказывает дополнительное костимулирующее влияние на пролиферацию Т-лимфоцитов.
· Активно пролиферирующие Т-клетки характеризуются нестабильностью генома, определяющей высокий риск появления дополнительных мутаций.
· В случае, если такая мутация имеет место, а мутировавшая клетка не элиминируется, продолжая активно пролиферировать, происходит образование ее многочисленных потомков с измененным геномом, т.е. моноклона Т-клеточной лейкемии.
· Таким образом, моноклональная Т-клеточная лейкемия/лимфома есть результат пролиферации одной единственной Т-клетки, подвергшейся дополнительной мутации.
Дата добавления: 2015-02-05; просмотров: 1493;