Пирамидная система.
Пирамидная система.Согласно традиционной точке зрения это основной механизм, реализующий
произвольные движения; начинается от моторных клеток Беца, находящихся в V слое моторной коры (4-е
поле), продолжается в виде корково-спинномозгового, или пирамидного, тракта, который переходит на
противоположную сторону в области пирамид и заканчивается на мотонейронах спинного мозга (на 2-м
нейроне пирамидного пути), иннервирующих соответствующую группу мышц.
К этим представлениям о пирамидной системе как об основном эфферентном механизме произвольных
движений добавились новые данные современной анатомии и физиологии.
Во-первых, не только 4-е поле является моторным. Это первичное моторное поле коры, разные участки
которого связаны с иннервацией различных групп мышц (см. схему «двигательного человечка» У.
Пенфилда и Г. Джаспера на рис. 11).
Как известно, первичное моторное поле коры больших полушарий характеризуется мощным развитием V
слоя, содержащего самые крупные клетки головного мозга человека (клетки Беца). Эти клетки («гигантские
пирамиды») обладают специфическим строением и имеют самый длинный аксон в нервной системе
человека (его длина может достигать двух метров), заканчивающийся на мотонейронах спинного мозга.
Моторные клетки пирамидного типа обнаружены не только в 4-м поле, но и в 6-м и 8-м полях
прецентральной коры, и во 2, 1 и даже в 3-м полях постцентральной коры (и в ряде других областей коры).
Следовательно, пирамидный путь начинается не только от 4-го поля, как это пред-
полагалось ранее, а со значительно больших площадей коры больших полушарий. По данным П. Дууса (1997), только 40 % всех волокон пирамидного пути начинается в 4-м поле, около 20 % — в постцентральной извилине; остальные — в премоторной зоне коры больших полушарий. Раздражение 4-го поля вызывает сокращение соответствующих групп мышц на противоположной стороне тела. Иными словами, 4-е поле построено по соматотопическому принципу.
Во-вторых, установлено, что пирамидный путь содержит волокна различного типа (по диаметру и степени миелинизации). Хорошо миелинизированные волокна составляют не более 10 % всех пирамидных волокон, которые идут от коры к периферии. По-видимому, с их помощью осуществляется фазический (непосредственно исполнительный) компонент произвольных движений. Подавляющее большинство слабо миелинизированных волокон пирамидного пути имеет, вероятно, иные функции и регулирует прежде всего тонические (фоновые, настроечные) компоненты произвольных движений.
В-третьих, если ранее предполагалось, что существует единый пирамидный, или кортико-спинальный, путь (латеральный), который идет с перекрестом в зоне пирамид от коры больших полушарий к мотонейронам спинного мозга, то в настоящее время выделен другой кортико-спинальный путь (вентральный), идущий без перекреста в составе пирамид на той же стороне. Эти два пути имеют различное функциональное значение. Наконец, пирамидный путь оканчивается не непосредственно на мотонейронах, расположенных в передних рогах спинного мозга, как считалось ранее, а главным образом на промежуточных (или вставочных) нейронах, с помощью которых модулируется возбудимость основных мотонейронов и тем самым оказывается воздействие на конечный результат — произвольные движения.
Все эти данные свидетельствуют о сложности пирамидной системы как исполнительного механизма (рис. 31).
Следует отметить, что кроме 4-го моторного поля (о моторных функциях которого известно уже очень давно, после опытов Г. Фритча и Е. Гитцига, раздражавших у животных электрическим током эту зону мозга) у человека обнаружен еще целый ряд моторных зон, при стимуляции которых также возникают двигательные эффекты. Эти зоны, получившие название дополнительных моторных зон, описаны Г. Джаспером, У. Пенфилдом и другими крупнейшими физиологами.
Хомская Е. Д. Х = Нейропсихология: 4-е издание. — СПб.: Питер, 2005. — 496 с: ил. 101
Существуют две основные дополнительные моторные зоны коры. Одна из них находится вдоль края Сильвиевой борозды; ее стимуля-
Рис. 31. Строение пирамидной и экстрапирамидной систем:
FG—NG—ML—VP—SP — проводящие пути и переключения кожно-кинестетических рецепций; CS—BS—NR — связи коры мозга с мозжечком; CR—MN — корково-двигательный пирамидный путь; CR—NR—RTS — связи коры с ядрами ретикулярной формации; CR—SNR — связи коры с черной субстанцией; Str— подкорковые узлы больших полушарий (по Д.
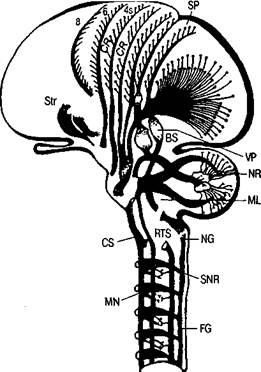
ция вызывает движения рук и ног (как ипсилатеральных, так и контралатеральных). Другая расположена на внутренней медиальной поверхности полушарий кпереди от моторной зоны, в прецентральных отделах мозга. Раздражение этой зоны также приводит к различным двигательным актам.
Важно отметить, что при раздражении дополнительных моторных зон возникают не элементарные сокращения отдельных мышечных групп, а целостные комплексные движения, что говорит об их особом функциональном значении (У. Пенфилд, Г. Джаспер, 1958).
Описаны и другие дополнительные моторные зоны коры. Все эти данные говорят о том, что современные
знания о корковой организации произвольных движений еще далеко не полны.
Помимо моторных зон коры больших полушарий, раздражение которых вызывает движения, существуют и
такие зоны коры, раздражение которых прекращает уже начавшееся движение (подавляющие области
коры). Они расположены кпереди от 4-го поля (поле 4s) на границе 4-го и 6-го полей; кпереди от 8-го поля
(поле 8s); кзади от 2-го поля (поле 2s) и кпереди от 19-го поля (поле 19s). На внутренней поверхности
полушария находится подавляющее поле 24s (см. рис. 4).
Поля 8s, 4s, 2s и 19s — это узенькие полоски, разграничивающие основные области коры, связанные с
проекциями ядер зрительного бугра на кору больших полушарий. Как известно, задние отделы коры (17, 18,
19-е поля) являются зоной проекции латерального коленчатого тела; сенсомоторная область — зона
проекции вентральных таламических ядер; префронтальная область — зона проекции ДМ-ядра зрительного
бугра. Таким образом, подавляющие полоски коры разграничивают сферы влияния разных реле-ядер
таламуса.
В коре больших полушарий имеются еще особые адверзивные зоны. Это области коры, хорошо известные
нейрохирургам и невропатологам. Их раздражение (электрическим током или болезненным процессом)
вызывает адверзивные эпилептические припадки (начинающиеся с адверзии — поворота туловища, глаз,
головы, рук и ног в сторону, противоположную расположению возбуждающего агента). Эпилепсия,
протекающая с припадками этого типа, известна как «эпилепсия Джексона».
Существуют две адверзивные зоны коры: премоторная и теменно-затылочная (поля 6, 8 и 19-е на границе с
37-м, 39-м полями). Предполагается, что эти поля коры участвуют в сложных реакциях, связанных с
вниманием к раздражителю, т. е. в организации сложных двигательных актов, опосредующих внимание к
определенному стимулу.
Хомская Е. Д. Х = Нейропсихология: 4-е издание. — СПб.: Питер, 2005. — 496 с: ил. 102
Дата добавления: 2014-12-11; просмотров: 1458;