Синаптическая передача
Синапс. Синапсом называют область соединения между окончанием аксона одного нейрона и дендритами или телом другого. Каждый нейрон может образовать до 800-1000 синапсов с другими нервными клетками, а плотность этих контактов в сером веществе мозга составляет боле 600 млн. на 1 ми3 (рис. A.30)1.
Место перехода нервного импульса с одного нейрона на другой
| Синаптические пузырьки |
| Синаптическая щель Пресинаптическа мембрана Постсинаптическ мембрана Митохондрии |
| Дендрит |
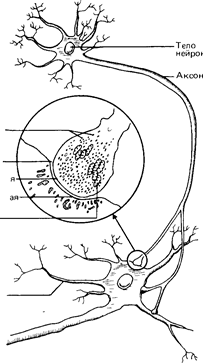
Рис. A.30. Синаптическое соединение нейронов (в середине - область синапса при большем увеличении). Терминальная бляшка пресинаптического нейрона содержит пузырьки с запасом нейромедиатора и митохондрии, доставляющие энергию. необходимую для передачи нервного сигнала.
1 Это значит, что если за одну секунду отсчитывать по 1000 синапсов, то для их полного пересчета потребуется от 3 до 30 тысяч лет (Changeux, 1983, р. 75).
Биологические основы поведения
| Молекулы Х Рецепторные ки |
| Нейрон 1 Нейрон Молекулы 1 а медиатора А |
| "Подходящие" молекулы психотрогжого препарата |
|
| Молекулы /^—модулятора R <?Л^ |
| 2а |

|

|

|
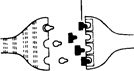
Рис. А.31. la. Медиатор А, молекулы которого освобождаются из концевой бляшки нейрона I, связывается специфическими рецепторами на дендритах нейрона II. МолекулыX, которые по своей конфигурации не подходят к этим рецепторам, занять их не могут и потому не вызывают каких-либо синаптических эффектов.
16. Молекулы М (например, молекулы некоторых психотропных препаратов) сходны по своей конфигурации с молекулами нейромедиатора А и поэтому могут связываться с рецепторами для этого медиатора, таким образом мешая ему выполнять свои функции. Например, ЛСД мешает серотонину подавлять проведение сенсорных сигналов.
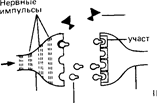
2а и 26. Некоторые вещества, называемые нейромодуляторами, способны воздействовать на окончание аксона, облегчая или подавляя высвобождение нейромедиатора.
представляет собой, собственно, не точку контакта, а скорее узкий промежуток, называемый синоптической щелью. Речь идет о щели шириной от 20 до 50 нанометров (миллионных долей миллиметра), которая с одной стороны ограничена мембраной пресинаптической бляшки нейрона, передающего импульс, а с другой - постсинаптической мембраной дендрита или тела другого нейрона, принимающего нервный сигнал и загем передающего его дальше.
Нейромедиаторы. Именно в синапсах происходят процессы, в результате которых химические вещества, освобождаемые пресинаптической мембраной, передают нервный сигнал с одного нейрона на другой. Эги вещества, получившие название нейромедиаторов (или просто медиато-ров),- своего рода «мозговые гормоны» (нейрогормоны) - накапливаюг-
268 Приложение А
ся в пузырьках синаптических бляшек и освобождаются, когда по аксону сюда приходит нервный импульс.
После этого медиаторы диффундируют в синаптическую щель и присоединяются к специфическим рецепторным участкам постсинаптиче-ской мембраны, т. е. к таким участкам, к которым они «подходят, как ключ к замку». В результате этого проницаемость постсинаптической , мембраны изменяется, и таким образом сигнал передается с одного нейрона на другой; медиаторы могут также и блокировать передачу нервных сигналов на уровне синапса, уменьшая возбудимость постси-наптического нейрона.
Выполнив свою функцию, медиаторы расщепляются или нейтрализуются ферментами либо всасываются обратно в пресинаптическое окончание, что приводит к восстановлению их запаса в пузырьках к моменту прихода следующего импульса (рис. А.31).
Возбуждающая или тормозная функция синапса зависит главным образом от типа выделяемого им медиатора и от действия последнего на постсинаптическую мембрану. Некоторые медиаторы всегда оказывают только возбуждающее действие, другие - только тормозное (инги-бирующее), а третьи в одних отделах нервной системы играют роль активаторов, а в других-ингибиторов.
Функции главных нейромедиаторов. В настоящее время известно несколько десятков этих нейрогормонов, но их функции изучены пока недостаточно. Сказанное, например, относится к ацетилхолину, который участвует в мышечном сокращении, вызывает замедление сердечного и дыхательного ритма и инактивируется ферментом ацетилхолинэстера-зoй\. Не вполне изучены и функции таких веществ из группы моноами-нов, как норадреналин, отвечающий за бодрствование мозговой коры и учащение сердечного ритма, дофамин, присутствующий в «центрах удовольствия» лимбической системы и некоторых ядрах ретикулярной формации, где он участвует в процессах избирательного внимания, или серотонин, который регулирует сон и определяет объем информации, циркулирующей в сенсорных путях. Частичная инактивация моноаминов происходит в результате их окисления ферментом моноаминоксидазой. Этот процесс, обычно возвращающий активность мозга к нормальному уровню, в некоторых случаях может приводить к чрезмерному ее снижению, что в психологическом плане проявляется у человека в чувстве подавленности (депрессии).
Гамма-аминомасляная кислота (ГАМК) представляет собой нейро-медиатор, выполняющий примерно ту же физиологическую функцию, что и моноаминоксидаза. Ее действие состоит главным образом в снижении возбудимостимозговых нейронов по отношению к нервным импульсам.
' По-видимому, недостаток ацетилхолина в некоторых ядрах промежуточного мозга-одна из главных причин болезни Альцгеймера. а недостаток до-фамина в скорлупе (одно из базальных ядер) может быть причиной болезни Паркинсона.
Биологические основы поведения 269
Наряду с нейромедиаторами существует группа так называемых нейромодуляторов, которые в основном участвуют в регуляции нервного ответа, взаимодействуя с медиаторами и видоизменяя их эффекты. В качестве примера можно назвать вещество Р и брадикинин, участвующие в передаче болевых сигналов. Освобождение этих веществ в синапсах спинного мозга, однако, может быть подавлено секрецией эндорфи-нов и энкефалина, которая таким образом приводит к уменьшению потока болевых нервных импульсов (рис. А.31, 2а). Функции модуляторов выполняют и такие вещества, как фактор S, играющий, по-видимому, важную роль в процессах сна, холецистокинин, ответственный за чувство сытости, ангиотензин, регулирующий жажду, и другие агенты.
Нейромедиаторы и действие психотропных веществ. В настоящее время известно, что различные психотропные препараты действуют на уровне синапсов и тех процессов, в которых участвуют нейромедиаторы и нейромодуляторы.
Молекулы этих препаратов по своей структуре сходны с молекулами определенных медиаторов, что и позволяет им «обманывать» различные механизмы синаптической передачи. Таким образом они нарушают действие истинных нейромедиаторов, либо занимая их место на рецеп-торных участках, либо мешая им всасываться обратно в пресинаптиче-ские окончания или подвергаться разрушению специфическими ферментами (рис. А. 31, 26).
Установлено, например, что ЛСД, занимая серотониновые рецептор-ные участки, мешает серотонину затормаживать приток сенсорных сигналов. Таким образом ЛСД открывает доступ к сознанию для самых разнообразных стимулов, непрерывно атакующих органы чувств.
Кокаин усиливает эффекты дофамина, занимая его место в рецептор-ных участках. Подобным же образом действуют морфин и другие опиаты, мгновенный эффект которых объясняется тем, что они быстро успевают занять рецепторные участки для эндорфинов1.
Действие амфетаминов обусловлено тем, что они подавляют обратное поглощение норадреналина пресинаптическими окончаниями. В результате накопление избыточного количества нейрогормона в синаптической щели приводит к чрезмерной степени бодрствования мозговой коры.
Принято считать, что эффекты так называемых транквилизаторов (например, валиума) объясняются главным образом их облегчающим влиянием на действие ГАМК в лимбической системе, что приводит к усилению тормозных эффектов этого медиатора. Наоборот, как антидепрессанты действуют главным образом ферменты, инактивирую-Щие ГАМК, или такие препараты, как, например, ингибиторы моноами-
Несчастные случаи, связанные с передозировкой наркотиков, объясняются тем, что связывание чрезмерного количества, например, героина эндорфиновыми рецепторами в нервных центрах продолговатого мозга приводит к резкому угнетению дыхания, а иногда и к полной его остановке (Besson, 1988, Science et Vie, Hors serie, n" 162).
270 Приложение А
ноксидизы, введение которых увеличивает количества моноаминов в синапсах.
Смерч ь от некоторых отравляющих газов наступает вследс1вие удушья. Такое действие этих газов связано с тем, что их молекулы блокируют секрецию фермента, разрушающего ацетилхолин. Между тем ацетилхолин вызывает сокращение мышц и замедление сердечного и дыхательного ритма. Поэгому его накопление в синаптических пространствах приводит к угнетению, а затем и полной блокаде сердечной и дыхательной функций и одновременному повышению тонуса всей мускулатуры.
Изучение нейромедиаторов еще только начинается, и можно ожидать, что в скором времени будут открыты сотни, а можег быть и тысячи этих веществ, многообразные функции которых определяют их первостепенную роль в регуляции поведения.
Дополнение А.4. Нервная активность и сканер
До недавнего времени единственным методом, позволяющим регистрировать электрическую активность мозга с помощью электродов, размещенных в разных участках черепной коробки, была электроэнцефалография (см. документ 4.1). Но записи, которые получают этим методом, с трудом поддаются расшифровке, и поэтому чаще всего электроэнцефалография дает лишь грубое представление об активности популяции нейронов, расположенных под электродом.
Недавно, однако, появилось другое устройство для регистрации нервной активности. Речь идет о так называемом сканере, позволяющем составлять довольно точные карты нервной активности в различных областях головного мозга.
Это устройство осуществляет томографическое сканирование головного мозга с помощью позитронной эмиссии (откуда и другое название сканера-позитронно-эмиссионный томограф). В основе метода лежит то обстоятельство, что для работы мозга используется главным образом глюкоза: чем выше активность данного участка, тем больше глюкозы ему требуется для поддержания работы.
Первый из такого рода методов заключается в выявлении активных зон мозга после инъекции в кровь радиоактивных изотопов (например. фтора-18 или углерода-11). способных испускать положительно заряженные частицы, называемые позитронами. Столкновение позитронов с отрицательно заряженными электронами в нейронах сопровождается «взрывом», в результате которого образуются два разлетающихся в противоположных направлениях фотона (см. документ 5.2). Эти кванты света, число которых должно быть больше в усиленно снабжаемых кровью активных участках, улавливались затем камерой с фоточувствительными элементами, производившей таким образом послойный анализ головного мозга. После определения компьютером точки возникновения каждого «взрыва» информация-точка за точкой -выводи
ло югические основы поведения 271

|
Рис. А.32. Позитронно-эмиссионный томограф (ПЭТ) благодаря камере с фоточувствительными элементами и компьютеру, интегрирующему данные, позволяет следить за распределением акчивности в головном мозгу и воспроизводить получаемую картину на экране
лась на телевизионный экран с изображением последовательных срезов
мозга (рис. А.32).
Однако то обстоятельство, что активность нейронов приходилось оценивать косвенным образом-по притоку крови, сильно снижало точность результатов. Недавно был предложен метод прямого определения активности нейронов с помощью внутриклеточной метки. Такая возможность появилась, когда было найдено вещество, которое клетка поглощает вместо глюкозы, но не может использовать. В результате это вещество -дезоксиглюкоза- способно накапливаться в нейронах и благодаря радиоактивной метке указывать места повышенной нервной активности. Поскольку, однако, позитрон за то время, которое проходит между его эмиссией и «взрывом» при столкновении с отрицательно заряженным электроном, успевает пробежать несколько миллиметров, получаемые изображения все-таки остаются несколько неточными. Тем не менее можно надеяться на быстрое усовершенствование описанного
Рис. А.33. Картины активности головного мозга, полученные с помощью ПЭТ (более активные участки выглядят более темными).
А. Активность в затылочных долях в зависимости от зрительного восприятия. С ieea: глаза испытуемого закрыты. В середине: испытуемый открывает глаза и видит однообразный белый фон. Справа: испытуемый видит перед собой парк. (По Phelps et al., 1982).

|
Б. Активность в височных и лобных долях во время прослушивания испытуемым рассказа о Шерлоке Холмсе (в середине) и «Бранденбургс-кого концерта» И.С.Баха (справа) С ieea: мозг испытуемого с заткнутыми ушами.
272 Приложение А
метода или на разработку других методов, которые позволят точнее картировать активность в мозгу.
Тем временем существующие методы уже дали возможность показать, что для бодрствования, например, характерна более высокая нервная активность в лобной доле, а эффекты сенсорной стимуляции выражены в тех участках мозга, которые имеют к данной сенсорной модальности более тесное отношение, чем к другим модальностям (Mazziota et al, 1982) (рис. А.ЗЗ).
Разумеется, в изучении нервной активности сделаны только первые шаги. Но, как полагает Шанжё, настанет, может быть, день, когда на телевизионном экране появится картина, воспроизводящая мысленный образ.
Резюме
Дата добавления: 2014-12-09; просмотров: 1160;