Глава 1. ВОДА-СРЕДА ОБИТАНИЯ РЫБ
Вода является средой, в которой зародилась органическая жизнь на Земле. Клеточная протоплазма первичных организмов по своим физико-химическим свойствам практически не отличалась от окружающей среды. С развитием эволюции усложнялось строение клеточной мембраны, что придавало цитозольной части клетки все большую независимость от внешней среды. У многоклеточных животных появились специализированные ткани: кожа, слизистые оболочки, которые сделали независимость внутренних процессов еще более совершенной. Однако полной победы над водной зависимостью нет ни у одного животного организма с планеты Земля, так как все внутренние процессы животных протекают в водных растворах и все животные организмы нуждаются в обновлении растворителя.
Вода выполняет важнейшую структурную роль в организме рыбы. Молекула Н2О с асимметрией в расположении атомов имеет вид биполярной конструкции. При контакте молекулы воды с липидной мембраной образуются бимолекулярные липидные мембраны. Вода образует гидратные оболочки и с белками. Интересно, что чем больше жизненный цикл животных привязан к водной среде, тем выше концентрация воды в тканях этих животных (табл. 1.1).
1.1. Содержание воды в теле животных, %
| Животные | Содержание воды, % | Животные | Содержание воды, % |
| Рыбы (карп) | Птицы (куры) | ||
| сеголетки | 89—75 | цыплята в возрасте 1—3 дня | 80—75 |
| взрослые | 70-65. | бройлеры товарные | 70—65 |
| ЗемноводныеRana sp | взрослые куры | 65—60 | |
| головастики | 90—80 | Млекопитающие (среднее) | |
| молодь массой 1-10 г | 76-70 | новорожденные | 80—70 |
| взрослые массой 150-200 г | 70-65 | взрослые | 60—50 |
Для представителей класса рыб вода является и основным по количеству компонентом клетки и остается внешней средой обитания.
Эволюционный прогресс связан с выходом животных из воды на сушу. Филогенетическую связь наземных животных с водной средой подчеркивает эмбриональное развитие птиц и млекопитающих. Эмбриогенез даже высших млекопитающих вызывает прямые ассоциации с жизнью в воде. На земле животные получили новую экологическую нишу, а эволюция в целом получила полигон для экспериментов по созданию новых видов. В этом свете ихтиофауна, остающаяся в водной среде, представляет эволюционные задворки. Рыбы обречены на вымирание по мере остывания Солнца и Земли, в то время как наземные обитатели получили шанс продлить свое присутствие на планете или за ее пределами.
Жизнь в водной среде накладывает отпечаток на морфологии и физиологию животных. Сопоставление физико-химических свойств водной и воздушной сред позволяет говорить и о преимуществах, и о недостатках водного образа жизни.
Вода - более плотная среда, чем воздух. Поэтому движение в воде более проблематично, чем в воздухе. Вместе с тем с плотностью воды связана нейтральная плавучесть водных животных и их способность легко менять положение тела в трехмерном пространстве. Гравитационное влияние на физическое тело в воде уменьшается, от чего в воде облегчается перемещение.
Вода, обладающая высокой теплоемкостью, нивелирует суточные и сезонные температурные перепады. Количественная характеристика энергетики водородных связей меняется в зависимости от температуры. Так, при 0њС вода утрачивает примерно 15 % водородных связей, а при 40 њС -уже 50%. Эта особенность води крайне важна для рыб. Образование и разрушение водородных связей сопровождаются выделением или поглощением тепловом энергии. Следовательно, при суточных и сезонных колебаниях температуры воздуха вода обеспечивает некоторую стабильности внешней и, что еще более существенно, внутренней среде организма рыб. Поэтому явление пойкилотермии обеспечивает в воде довольно равномерную физиологическую активность в течение года, тогда как наземная пойкилотермия сопровождается такими явлениями, как зимняя спячка, ночное оцепенение.
На заре формирования животного мира на Земле пойкилотермия была естественным и единственно оправданным явлением. Температура воды первичного океана была благоприятна для биохимических процессов, а огромная масса океана гарантировала относительное температурное постоянство. В изменяющихся в историческом смысле условиях жизни на Земле больше шансов выжить у видов, защищенных от температурных перепадов. Поэтому гомойотермия рассматривается биологами как крупнейший ароморфоз, т. е. как важнейшее эволюционное приобретение.
Довольно велики различия в свойствах воды и воздуха в плане обеспечения животных кислородом. Здесь следует признать несомненное преимущество воздуха как дыхательной среды. Растворимость кислорода в воде невелика - максимум 15 мг/л при нормальных атмосферных условиях. В таком же объеме воздуха содержится в 20 раз больше кислорода. Надо отметить и то, что извлечь животным кислород из воды значительно труднее. Чтобы в организм рыбы поступило необходимое количество кислорода, ей приходится прокачивать через органы дыхания большую массу воды. Кроме того, дыхание у рыб осложняется и тем, что жабры как орган помимо дыхания выполняют и другие функции, например осморегуляцию, кроветворение, регуляцию кровообращения.
Однако благодаря противоточному характеру движения воды и крови через жабры эффективность извлечения кислорода из воды очень высока. Интересно отметить, что жабры являются очень хорошим теплообменником, который обеспечивает терморегуляцию. Жабры обеспечивают равенство температуры окружающей воды и внутренней среды организма рыб. В животном мире гомойотермия появляется лишь после утраты животными в процессе эволюции жаберного аппарата.
Солевой состав воды также создает определенные проблемы для рыб. Содержание воды в организме рыб составляет 60-80 %, содержание минеральных веществ -0,6-1 %. У морских рыб и миксин солевой состав крови практически не отличается от солевого состава среды их обитания. Поэтому у них нет серьезного осмотического барьера. И в пресной воде морские виды очень быстро погибают из-за нарушения водно-солевого состава тела. У пресноводных и проходных рыб механизм осморегуляции очень эффективен и заслуживает отдельного рассмотрения.
§ 1. ОСМОРЕГУЛЯЦИЯ РЫБ
Все виды животных имеют механизм осморегуляции. Осмотическое давление клетки -это основа мембранного потенциала, определяющего важнейшие свойства тканей - раздражимость и возбудимость. Осмотическое давление можно рассматривать как первый признак настоящей животной клетки, зародившейся в первичном океане.
У рыб можно выделить два основных механизма осморегуляции.
Первый (примитивный) механизм осморегуляции характерен Для акул. Плазма, лимфа и другие жидкости организма этих рыб по электролитному составу мало отличаются от морской воды, соленость которой равнозначна I % NaCl. На первый взгляд в таких условиях отпадает необходимость тратить энергию на осморегуляцию. Однако специфика азотистого обмена такова, что в крови акул накапливается большое количество (5 %) мочевины и триметиламиноксида, которые плохо диффундируют через мембраны клеток почек и жабр. Поэтому у акул возникает повышенное осмотическое давление внутренней среды (26атм против 24атм в морской воде) (1атм= 105Па).
Это создает условия для свободного проникновения воды из внешней среды в межклеточную жидкость и кровь,; что, в свою очередь, меняет градиент концентрации NaCl и приводит к диффузии натрия и калия. Фактически мембраны удерживают только мочевину.
Организм акулы представляет собой устойчивую биологическую систему, т. е. обладает гомеостазом, как и у всех других животных. Другими словами, у акулы есть механизм поддержания электролитного состава. Эволюция снабдила акул для поддержания гомеостаза специальным органом - ректальной железой, которая выводит в клоаку избыток солей.
Второй тип осморегуляции присущ костистым рыбам. Этот механизм более эффективен, однако и более сложен (рис. 1.1). Костистые обитают как в пресной, так и в соленой воде. Поэтому и внутри этой группы есть некоторые различия в механизме осморегуляции. У пресноводных костистых рыб осмоляльность крови выше, чем окружающей их воды: соответственно 6 и 0,3 атм. В соленой воде картина обратная: осмотическое
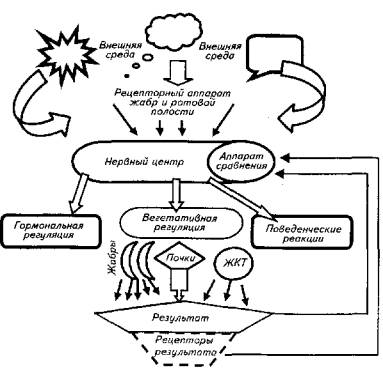
Рис. 1.1. Схема функциональной системы осморегуляции рыб
давление крови составляет 9 атм, а морской воды - 24 атм. Следовательно, и в первом, и во втором случае требуются затраты энергии для поддержания осмотического давления внутренней среды организма рыб.
В морской воде покровные ткани костистых рыб избирательно пропускают воду и не пропускают растворенные в ней соли. Рыбы активно заглатывают воду, однако слизистая желудочно-кишечного тракта (ЖКТ) избирательно абсорбирует только воду и одновалентные ионы натрия, калия, хлора. Остальные удаляются из организма в составе каловых масс. Вероятный избыток двухвалентных ионов (кальций, магний, сульфат) выводится почками. Помимо этого имеется и дополнительный механизм регуляции ионного состава.
Слизистая жаберного аппарата включает в себя сеть специфических клеток, которые экскретируют во внешнюю среду ионы хлора и натрия. Они так и называются - "хлоридные". Необходимо подчеркнуть, что жабры имеют особый мембранный потенциал, который облегчает экскрецию катионов. Этот потенциал оценивается в 20-25 мВ. Учитывая, что снаружи жаберная мембрана несет положительный заряд, экскреция натрия в таких условиях становится довольно эффективной даже без затрат энергии.
В пресной воде угрозу гомеостазу создает сама вода (не электролиты). Поэтому заглатывание воды пресноводными рыбами биологически нецелесообразно. Однако через жабры вода диффундирует активно. Компенсируется эта избыточность очень эффективной работой почек. У костистых рыб в пресной воде образуются и выделяются сравнительно большие количества мочи. По крайней мере, эта величина на порядок выше, чем у морских пластинчатожаберных. Почки пресноводных имеют очень высокую эффективность реабсорбции электролитов.
Солевой гомеостаз регулируется нейрогуморальным путем. Осморецепторы располагаются не только в кровеносных сосудах, но и на слизистых жабр и ротовой полости. Их возбуждение приводит к активизации гипоталамо-гипофизарного механизма. Непосредственно на проницаемость мембран, через которые осуществляется транспортирование электролитов и воды, влияют соматотропин, пролактин, вазопрессин, окситоцин, кортизол. Помимо проницаемости мембран эти гормоны контролируют синтез белков - переносчиков ионов, влияют на регуляцию кровотока, Фильтрации и реабсорбции в почках.
Жаберные мембраны более активно пропускают электролиты за счет активного переноса и диффузии по сравнению с водой (только диффузия). Мембранный потенциал жаберных мембран костистых рыб в пресной воде составляет минус 30 - минус 40 мВ, что благоприятствует переносу катионов из внешней среды в кровь.
Осморегуляция находится в большой зависимости и от функционального состояния органов дыхания. Так, гипоксия и сопровождающая ее одышка неизбежно приводят к поступлению в кровь избыточных количеств воды через жабры. Экспериментальная гипоксия мальков лосося приводит к увеличению живой массы на 15 % за счет обводнения.
Несмотря на наличие специфических органов осморегуляции, основная работа в этом процессе выполняется почками. Поэтому именно здесь представляется целесообразным рассмотреть механизм мочеобразования рыб.
§ 2. ПОЧКИ КАК ОРГАН ОСМОРЕГУЛЯЦИИ
Почки особенно хорошо развиты у костистых пресноводных рыб. Подсчитано, что у взрослой форели величина диуреза колеблется от 200 до 300 мл на 1 кг живой массы. Диурез в морской воде сокращается в 10 раз. Именно в этих пределах изменяется интенсивность диуреза у эвригалинных видов костистых рыб при их миграциях. В механизме адаптации рыб центральное место занимает эндокринная система.
В классе рыб отмечается большое разнообразие морфологию почек. Самые примитивные почки у рыбообразной миксины. Почка у нее представляет собой воронкообразное скопление клеток реснитчатого эпителия в брюшной полости, называемое нефростомом. Реснички нагнетают в воронку полостную жидкость, где и происходят фильтрация и реабсорбция. Понятно, что нефростом работает малоэффективно.
У более совершенных видов рыб почки устроены сложнее. Они представляют собой два тяжа паренхимы, расположенные вдоль тела параллельно спинной аорте. Проксимальная часть почек не имеет отношения к осморегуляции и отвечает за кроветворение, иммунитет, а также выполняет эндокринную функцию. Средняя и дистальная части почек обеспечивают осморегуляцию и выделение.
Структурным элементом почки является нефрон, который у рыб имеет аналогичное с другими позвоночными строение и состоит из сосудистого клубочка и капсулы с отходящим от нее мочесобирательным канальцем (рис. 1.2).
Базальная мембрана капсулы соприкасается с сосудистым клубочком. Из-за разницы осмотического давления вода, одно- и двухвалентные ионы легко диффундируют из капилляра в полость капсулы.
Образующаяся в капсуле жидкость и есть первичная моча. Ее состав близок к составу плазмы крови и не содержит лишь липиды и белки. В канальцах происходит обратное всасывание одновалентных ионов, аминокислот, мочевины и триметиламина. Здесь же реабсорбируется 70-80 % воды. В свою очередь, эпителиальные клетки мочесобирательного канальца секретируют двухвалентные ионы и креатинин.
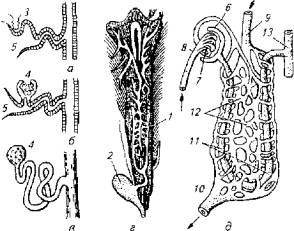
Рис. 1.2. Макро- и микростроение почек:
а- головная почка; б, в, д - туловищная почка; г -почка форели; 7 - мочеточники; 2- мочевой пузырь; 3- наружный клубочек; 4- мальпигиево тельце; 5- воронка; 6- капсула; 7- сосудистый клубочек; 8-спинная аорта; 9- воротная вена почек; 10- почечная вена; 11 - почечные канальцы; 12 - венозные синусоиды; 13 - собирательная трубка
В результате фильтрации плазмы крови в капсуле Боумена и процессов реабсорбции и секреции в канальцах формируется вторичная моча. В табл. 1.2 на примере форели показаны основные ионы мочи.
Дата добавления: 2014-12-22; просмотров: 2843;