Строение растительной клетки
Клетка – термин (от греческого cytos – «клетки» или латинского cellula – «полость») впервые употребил Роберт Гук в 1665 г. при описании строения пробки. Позже наблюдения Р. Гука повторили Грю, Мальпиги и другие исследователи на различных растениях. Однако признание универсальности клеточного строения всего живого произошло лишь в 1838–1839 гг., когда была сформулирована клеточная теория независимо друг от друга ботаником М. Шлейденом и зоологом Т. Шванном. Наука, занимающаяся микроскопическим изучением клетки, называлась в то время цитологией. В конце XIX в., а затем уже в ХХ в., изучение клеток приобрело в значительной мере экспериментальный характер, и теперь существует целая большая отрасль науки, именуемая биологией клетки, которая использует самые разнообразные методы для того, чтобы постичь жизнедеятельность организма на клеточном уровне.
Усовершенствование методов исследования и использование физических и химических подходов привело к успешному проникновению в тайны организации клетки. Было выявлено единство в строении клетки разных организаций, доказана связь между ее структурой и функцией. Основные положения клеточной теории, которые были сформулированы более 150 лет назад, были развиты и углубленны и на современном этапе развития биологии формулируются следующим образом:
1. Клетка является основной структурой и функциональной единицей жизни. Все организмы состоят из клеток, жизнь организма в целом обусловлена взаимодействием составляющих его клеток.
2. Клетки всех организмов сходны по химическому составу, строению и ряду функций.
3. Все новые клетки образуются при делении начальных клеток.
Для всех клеток характерна способность к росту, размножению, дыханию, выделению, использованию и преобразованию энергии, они реагируют на раздражение. Таким образом, клетки обладают всей совокупностью свойств, необходимых для поддержания жизни. Отдельные части клеток не могут выполнять весь комплекс жизненных функций, только совокупность структур, которые образуют клетку, проявляют все признаки жизни. Поэтому только клетка является основной структурой и функциональной единицей живых организмов. Клетка – это самостоятельная саморегулирующая химическая система.
В многоклеточных организмах, в том числе и в растениях, отдельные клетки тесно и слаженно взаимодействуют одна с другой.
Размеры клетки даже в пределах одного организма очень разные, что, в значительной мере, зависит от их специализации и выполняемых функций. Они могут быть в виде многогранников, могут иметь шаровидную, кубическую и другие формы.
Размеры клеток – от нескольких десятков микрометров до нескольких сантиметров: клетка с зоны растяжения стебля или корня имеет размеры 50×20×10 мкм, а клетка харовой водоросли – несколько сантиметров в длину и до 1 мм в диаметре.
Строение растительной клетки довольно сложное и высокодифференцированное, но, на первый взгляд, можно выделить три компартмента (три отдельных пространства): клеточная стенка, протоплазма и вакуоль.
Существует значительное количество классификаций структурных элементов клетки: рассмотрим одну из них (рис. 1.1).

|
| Рис. 1.1. Классификация структурных элементов растительной клетки |
Эта классификация несколько отличается от общепринятой. Обычно все компоненты клетки, за исключением ядра, обозначают общим понятием цитоплазма, причисляя митохондрии и пластиды к «органеллам цитоплазмы», хотя присутствие в них собственного материала позволяет в той же степени отделить их от цитоплазмы, как это принято при рассмотрении клеточного ядра. Рибосомы имеются не только в цитоплазме, но и в структурных элементах, в том числе в клеточном ядре.
Клеточная стенка, вакуоль и пластиды – типичные образования растительной клетки, не встречающиеся в клетках животных.
Клеточные стенки.Материал для построения клеточной стенки секретирует сам заключенный в ней – протопласт. Клеточная стенка, отлагающаяся во время деления клеток растения, называется первичной клеточной стенкой. Позже в результате утолщения она может превратиться во вторичную клеточную стенку.
Первичная клеточная стенка состоит из целлюлозных мицелл, которые образуют микрофибриллы, погруженных в матрикс, в состав которого входят сложные полисахариды.
 Целлюлоза представляет собой полисахарид (полимер глюкозы). В ней заключено около 50 % углерода, находящегося в растениях. 20–40 % материала клеточной стенки составляет целлюлоза. Целлюлозные волокна представляют собой длинные цепи – приблизительно из 10 000 остатков глюкозы. Из каждой цепи выступает наружу множество ОН-групп. Эти группы направлены во все стороны и образуют водородные связи с соседними цепями, что обеспечивает жесткое поперечное сшивание всех цепей. Цепи объединены друг с другом, образуя микрофибриллы, в которых происходит образование пространственных решеток, кристаллоподобных участков, так называемых мицелл (рис. 1.2).
Целлюлоза представляет собой полисахарид (полимер глюкозы). В ней заключено около 50 % углерода, находящегося в растениях. 20–40 % материала клеточной стенки составляет целлюлоза. Целлюлозные волокна представляют собой длинные цепи – приблизительно из 10 000 остатков глюкозы. Из каждой цепи выступает наружу множество ОН-групп. Эти группы направлены во все стороны и образуют водородные связи с соседними цепями, что обеспечивает жесткое поперечное сшивание всех цепей. Цепи объединены друг с другом, образуя микрофибриллы, в которых происходит образование пространственных решеток, кристаллоподобных участков, так называемых мицелл (рис. 1.2).
 |
Особо важное значение для той роли, которую выполняет целлюлоза в клеточных стенках имеет ее волокнистое строение и высокая прочность на разрыв, сравнимая с прочностью стали.
Микрофибриллы, погруженные в матрикс, образуют каркас клеточной стенки. Матрикс клеточной стенки состоит из полисахаридов, которые для удобства делят на пектины и гемицеллюлозы в зависимости от их растворимости в различных растворителях.
Гемицеллюлозы – это смешанная группа полисахаридов, растворимая в щелочах (к ним относятся полимеры ксилозы, галактозы, маннозы, глюкозы и глюкоманнозы). У гемицеллюлозы, как и у целлюлозы, молекулы имеют форму цепи, однако их цепи короче, менее упорядочены и сильнее разветвлены.
Молекулы пектина представляют собой цепочки полигалактуроновой кислоты, свободные карбоксильные группы которой составляют резервуар фиксированных отрицательных зарядов в клеточных стенках.
Отметим также, что размер межмицеллярного и межфибриллярного пространства (пор) составляет соответственно 1 и 10 нм, что значительно превышает размер многих растворенных веществ, транспортируемых в растениях.
Среди вещества матрикса клеточных стенок имеются и белки. Это гликопротеиды, белковая фракция которых сильно обогащена аминокислотой оксипролином. Структурный белок клеточной стенки получил название экстенсина.
Таким образом, свободные карбоксильные группы полисахаридов и белков обеспечивают большое число отрицательных фиксированных зарядов в клеточных стенках. Свободные аминогруппы белков ответственны за меньшее количество положительных зарядов. Заряженные группы матрикса образуют доннановскую систему, играющую важную роль в процессе транспорта ионов в тканях растений. Клеточные стенки гидратированы: 60–70 % их массы обычно составляет вода. По свободному пространству клеточной стенки вода перемещается беспрепятственно. Присутствие воды оказывает влияние на химические и физические свойства полисахаридов клеточной стенки.
У некоторых клеток, например у клеток мезофилла листа, на всем протяжении их жизни имеется только первичная клеточная стенка. Однако у большинства клеток на внутреннюю поверхность первичной стенки отлагаются дополнительные слои целлюлозы, т. е. образуется вторичная клеточная стенка. Обычно это происходит после того, как клетка достигнет своего максимального размера, и лишь немногие клетки, например клетки колленхимы, продолжают рост во время этой фазы. Вторичное утолщение клеточных стенок растения не следует путать со вторичным утолщением (вторичным ростом) самого растения, т. е. увеличением толщины ствола в результате добавления новых клеток.
Самый внутренний, очень тонкий последний слой вторичной стенки вследствие особенностей структуры и состава принято называть третичной стенкой. Наряду с целлюлозой и пектином в этом слое имеется много гемоцеллюлоз: в третичной стенке микрофибриллы, по-видимому, образуют сетчатую структуру. Обращенная внутрь клетки поверхность третичной стенки обычно покрыта субмикроскопическими бородавчатыми выростами.
Некоторые клетки, такие, как трахеальные элементы ксилемы и клетки склеренхимы, претерпевают интенсивную лигнификацию (одревеснение); при этом все слои целлюлозы (первичный и вторичные) пропитываются лигнином – сложным полимерным веществом, не относящимся к полисахаридам. В клетках протоксилемы отложения лигнина имеют кольцевую, спиральную или сетчатую форму. В других случаях лигнификация бывает сплошной, если не считать так называемых поровых полей, т. е. тех участков в первичной клеточной стенке, через которые осуществляется контакт между соседними клетками при помощи плазмодесм. Лигнин скрепляет целлюлозные волокна и удерживает их на месте. Он действует как очень твердый и жесткий матрикс, усиливающий прочность клеточных стенок на растяжение и в особенности на сжатие. Он же обеспечивает клеткам дополнительную защиту от неблагоприятных физических и химических воздействий. В ряде случаев клетки пропитываются суберином или покрыты восковым налетом – кутикулой.
Клеточная стенка, как упоминалось, пронизана порами, сквозь которые проходят плазмодесмы. Плазмодесма – это не просто нить цитоплазмы, соединяющая протопласты соседних клеток, а органелла имеющая сложное строение. Снаружи плазмодесма покрыта плазмалеммой, которая переходит из одной клетки в другую. Внутри плазмодесмы находится канал (десмотубола), соединяющий эндоплазматическую сеть соседних клеток. Пора, находящаяся в клеточной стенке, в средней части расширена, а на концах сужена, поэтому на концах плазмалемма почти вплотную подходит к десмотуболе. На 1 мм2 клеточной стенки приходится от 100 тыс. до 50 млн. плазмодесм.
Совокупность протопластов всех клеток, соединенных плазмодесмами, получила название симпласта.
Сравнительно недавно были открыты эктодесмы – очень тонкие цитоплазматические выросты, пронизывающие наружные стенки эпидермальных клеток листа и доходящие до кутикулы. Они подвижны: могут укорачиваться и удлиняться. Эктодесмы, вероятно, выполняют в многоклеточном организме роль рецепторов: воспринимают разного рода сигналы об изменениях в окружающей среде и передают их затем симпласту.
Основные функции клеточной стенки (оболочки) следующие:
1. Оболочки обеспечивают отдельным клеткам и растению в целом механическую прочность и опору. В некоторых тканях прочность усиливается благодаря интенсивной лигнификации клеточных стенок (небольшое количество лигнина присутствует во всех клеточных стенках).
2. Относительная жесткость клеточных стенок и сопротивление растяжению обуславливают и тургесцентностьклеток, когда в них осмотическим путем поступает вода. Это усиливает опорную функцию во всех растениях и служит единственным источником опоры для травянистых растений и для таких органов, как листья, т. е. там, где отсутствует вторичный рост. Клеточные стенки также предохраняют клетки от разрыва в гипотонической среде.
3. Ориентация целлюлозных микрофибрилл ограничивает, и в известной мере, регулирует как рост, так и форму клеток, поскольку от расположения этих микрофибрилл зависит способность клеток к растяжению. Если, например, микрофибриллы располагаются поперек клетки, окружая ее как бы обручами, то клетка, в которую путем осмоса поступает вода, будет растягиваться в продольном направлении.
4. Система связанных друг с другом клеточных стенок (апопласт) служит путем, по которому передвигается вода и минеральные вещества. Клеточные стенки скреплены между собой с помощью срединных пластинок. В стенках имеются небольшие поры, сквозь которые проходят цитоплазматические тяжи – плазмодесмы. Плазмодесмы объединяют все протопласты в единую систему – симпласт.
5. Наружные клеточные стенки эпидермальных клеток покрываются особой пленкой – кутикулой, состоящей из воскообразного вещества кутина, что снижает потерю воды и уменьшает риск проникновения в растение болезнетворных организмов. В пробковой ткани клеточные стенки по завершению вторичного роста пропитываются суберином, выполняющим сходную функцию.
6. Клеточные стенки сосудов ксилемы, трахеид и ситовидных трубок приспособлены для дальнего транспорта веществ по растению.
7. Стенки клеток эндодермы корня пропитаны суберином и поэтому служат барьером на пути движения воды и минеральных веществ.
8. У некоторых клеток их видоизмененные стенки хранят запасы питательных веществ. Таким способом, например, запасаются гемицеллюлозы в некоторых семенах.
9. У передаточных клеток (клетки-спутники) площадь поверхности клеточных стенок увеличена и соответственно увеличена площадь поверхности плазмалеммы, что повышает эффективность переноса веществ.
10. Благодаря своим катионобменным свойствам клеточная стенка служит резервуаром, накапливающим катионы.
Далее, все что находится за клеточной оболочкой, составляет протопласт.
Протопласт – гетерогенная система. Снаружи протопласт окружен мембраной – плазмалеммой. В наши дни существуют методы выделения протопластов из клеток. Для этого клетки помещают в высококонцентрированный раствор какого-либо осмотически активного вещества, содержащий, кроме того, ферменты, способные разрушать клеточную стенку.
Химический состав протопластов самых различных тканей может быть охарактеризован следующими цифрами: 75–85 % воды, 10–20 % белка, 2–3 % липидов, 1 % углеводов и около 1 % солей.
В соответствии с приведенной классификацией нужно выделить протоплазму и вакуоль. В состав протоплазмы входят структуры, какие указаны на рис. 1.1.
Цитоплазма – компартмент, в котором происходят основные процессы жизнедеятельности клетки. Цитоплазма состоит из водянистого вещества и находящихся в нем разнообразных органелл. Цитозолем (гиалоплазма) называют растворимую часть цитоплазмы – это основное вещество, заполняющее пространство между клеточными органеллами. Цитозоль содержит систему микрофиламентов. На долю воды в цитоплазме приходится приблизительно 90 %, в которой, в растворимой форме содержатся все основные биомолекулы. Истинный раствор образуют ионы и малые молекулы, а именно соли, сахара, аминокислоты, жирные кислоты, нуклеотиды, витамины и растворенные газы. Крупные молекулы – белки и в меньшей мере газы. Крупные молекулы – белки и в меньшей мере РНК – образуют коллоидные растворы. Коллоидный раствор может быть золем (невязким) или гелем (вязким). Цитозоль – это не только место хранения биомолекул. Здесь протекают и метаболические процессы, среди них такой важный процесс, как гликолиз. Синтез жирных кислот, нуклеотидов и некоторых аминокислот также происходит в цитозоле.
В обычных условиях цитоплазма находится в активном состоянии: заметно движение органелл, а в ряде случаев всей цитоплазмы – циклоз.
Вакуоли отделены от цитоплазмы тонопластом. Жидкость, заполняющая вакуоль представляет собой концентрированный водный раствор, содержащий минеральные соли, сахара, органические кислоты, кислород, диоксид углерода, пигменты и некоторые отходы жизнедеятельности или вторичные продукты метаболизма.
Согласно принятой в настоящее время точке зрения, вакуоли могут возникать только из предсуществовавших уже вакуолей или из каких-то других полостей клетки, ограниченных мембранами, например при разрастании пузыревидных выпячиваний эндоплазматической сети или увеличении пузырьков Гольджи.
Вакуоли выполняют следующие функции:
1. Вода обычно поступает в концентрированный вакуолярный сок путем осмоса через избирательно проницаемый тонопласт. В результате в клетке развивается тургорное давление, и цитоплазма прижимается к клеточной стенке. Осмотическое поглощение воды играет важную роль при растяжении клеток во время роста, а также в общем водном режиме растения.
2. Иногда в вакуоле присутствуют пигменты, называемые антоцианами. В эту группу входят антоцианианы, имеющие красную, синею или пурпурную окраску, и некоторые родственные соединения окрашенные в желтый или кремовый цвета. Именно эти пигменты главным образом и определяют окраску цветов (например, у роз, фиалок, георгинов), а также окраску плодов, почек и листьев. У последних они обуславливают различные оттенки осенью. Окраска играет роль в привлечении насекомых, птиц и некоторых других животных, участвующих в опылении растений и в распространении семян.
3. У растений в вакуолях содержатся иногда гидролитические ферменты, и тогда при жизни клетки вакуоли действуют как лизосомы. После гибели клетки тонопласт, как и все другие мембраны, теряет свою избирательную проницаемость, и ферменты высвобождаются из вакуолей, вызывая автолиз.
4. В вакуолях растения могут накапливаться отходы жизнедеятельности и некоторые вторичные метаболиты. Из отходов иногда обнаруживаются кристаллы оксалата кальция. Роль вторичных продуктов не всегда ясна. Это касается в частности алкалоидов, которые могут сохраняться в вакуолях. Возможно, что они, подобно танинам с их вяжущим вкусом, отталкивают травоядных животных, т. е. выполняют защитную функцию. Танины особенно часто встречаются в вакуолях (как впрочем, и в цитоплазме, и в клеточных стенках) листьев, коры, древесины, незрелых плодов и семенных оболочках. Может накапливаться в вакуолях и латекс (млечный сок растений), обычно в виде молочно-белой эмульсии, например, млечный сок одуванчика.
Отдельные клетки (млечные клетки) специализируются на выделении млечного сока. Так в млечном соке бразильской гевеи содержаться ферменты и соединения, необходимые для синтеза каучука, а в млечном соке мака снотворного – алкалоиды.
5. Некоторые из компонентов вакуолярного сока играют роль запасных питательных веществ, при необходимости используемых цитоплазмой. Среди них в первую очередь следует назвать сахарозу, минеральные соли и инулин.
Ядра имеются во всех эукариотических клетках, за исключением зрелых члеников ситовидных трубок флоэмы и зрелых эритроцитов млекопитающихся и являются наиболее заметной структурой в цитоплазме клетки.
Ядро окружено двойной мембраной, которую часто называют ядерной оболочкой. В ядерной мембране имеются крупные, видимые в электронный микроскоп поры (диаметр 30–100 нМ; приблизительно 1 000 пор на одно ядро). Диаметр ядра составляет 5–10 мкм. В этом отношении ядерная мембрана отличается от всех других мембран. Ядерная пора – это непростое отверстие, а сложная структура, через которую проходят макромолекулы или даже рибосомы. Однако небольшие аминокислотные молекулы поступают в ядро не путем диффузии через поры, а путем активного транспорта через ядерную мембрану. Ядро необходимо для жизни клетки, поскольку именно оно регулирует всю активность. В клеточном ядре находятся ДНК-содержащие хромосомы и РНК-содержащие ядрышки, погруженные в свободный от нуклеиновых кислот матрикс, называемый нуклеоплазмой.
В форме ДНК в хромосомах содержится генетическая информация, с помощью которой контролируются все клеточные процессы обмена веществ, роста и развития.
Ядро выполняет следующие функции: 1) хранение информации; 2) передача информации от клетки к клетке (деление ядра, клеточное деление, размножение, наследственность) путем синтеза абсолютно идентичной ДНК, в которой закодирована эта информация; 3) передача информации в цитоплазму путем синтеза информационной РНК.
Нуклеоплазма, или ядерный сок – это внутренняя среда ядра, его матрикс. В матриксе есть зоны клеточного вещества – хроматина, который легко окрашиваются щелочными красителями. В состав хроматина входит ДНК и специальные белки – гистоны. В нуклеоплазме содержатся кислые белки (в противоположность основным гистонам хромосом), среди которых имеются многие ферменты
Каждое ядро содержит одно или несколько ядрышек, которые можно увидеть под световым микроскопом.
Ядрышки представляют собой округлые образования. В ядрышках синтезируются рибосомные РНК. Обычно в ядрах диплоидных организмов имеется два ядрышка, по одному для каждого гаплоидного набора хромосом. Состоят они в основном из белка, ядрышки содержат до 5 % РНК.
Эндоплазматическая сеть (эндоплазматический ретикулум) представляет собой систему связанных полостей, канальцев и трубочек, которые пронизывают всю мезоплазму (от плазмалеммы до тонопласта). Эти полости и каналы окружены мембраной.
К мембранам ЭР прикреплено большое количество рибосом – органоидов, которые состоят из белков и РНК приблизительно в одинаковых весовых соотношениях.
Рибосомы выявлены также в гиалоплазме, ядре, пластидах и митохондриях. Независимо от местонахождения выполняют одну и туже функцию – участвуют в синтезе белков.
Для характеристики молекулярной массы больших молекул, в частности рибосом, используют константы седиментации, которые измеряются с помощью ультроцентрофугирования. Константу седиментации выражают в единицах Сведберга (S), которые для эукариот составляют 80 S, прокариот – 70 S, для митохондрий и хлоропластов – 70 S.
Аппарат Гольджи – термин, который используется для обозначения всех диктиосом в клетке. Диктиосомы – это группы плоских, дисковидных пузырьков, или цистерн, которые по краям разветвляются в сложную систему трубочек. Диктиосомы в клетках высших растений обычно состоят из четырех – восьми цистерн, собранных вместе. Обычно в пачке цистерн различают формирующуюся и созревающие стороны. Мембраны, формирующихся цистерн по структуре напоминают мембраны ЭР, а мембраны созревающих цистерн – плазматическую мембрану.
Диктиосомы участвуют в секреции, а у большинства высших растений – в образовании клеточных оболочек. Полисахариды клеточной оболочки, синтезируемые диктиосомами, накапливаются в пузырьках, которые затем отделяются от созревающих цистерн. Эти секреторные пузырьки мигрируют и сливаются с плазматической мембраной, при этом содержащиеся в них полисахариды встраиваются в клеточную оболочку.
Лизосомы – это овальной формы органоиды клетки (пузырьки диаметром 1 мкм), окруженные мембраной. В них содержится набор ферментов, которые разрушают белки, нуклеиновые кислоты, углеводы, липиды. Ферменты лизосом (гидролитические) расщепляют принесенные пузырьками вещества.
Мембрана лизосомпрочная и затрудняет проникновение собственных ферментов в цитоплазму клетки, но когда лизосома повреждается какими-либо внешними факторами, то разрушается вся клетка или часть ее. Лизосомы обеспечивают дополнительным «сырьем» химические и энергетические процессы в клетке.
В типичной клетке имеется множество митохондрий – органелл, в которых происходит дыхание, а точнее синтез АТФ.
Митохондрии имеют округлую и продолговатую форму диаметром 0,4–0,5 мкм и длиной 1–5 мкм (рис. 1.3).
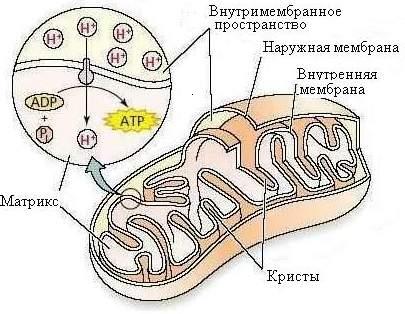
|
| Рис. 1.3. Строение митохондрии |
Количество митохондрий варьирует от единиц до 1 500–2 000 на растительную клетку.
Митохондрии ограничены двумя мембранами: наружной и внутренней, толщина каждой из них 5–6 нм. Наружная мембрана выглядит растянутой, а внутренняя образует складки, называемые гребнями (кристами), различной формы. Пространство между мембранами, в состав которого входит также внутреннее пространство крист, называется межмембранным (перимитохондриальным) пространством. Оно служит средой для внутренней мембраны и матрикса митохондрий.
Митохондрии в целом содержат 65–70 % белка, 25–30 % липидов и небольшое количество нуклеиновых кислот. 70 % от общего содержания липидов составляют фосфолипиды (фосфатидилхолин и фосфатидилэтаноламин). Жирнокислотный состав характеризуется высоким содержанием насыщенных жирных кислот, обеспечивающих «жесткость» мембраны.
В митохондриях локализованы системы аэробного дыхания и окислительного фосфорилирования. В результате дыхания расщепляются органические молекулы, и высвобождается энергия с передачей ее на молекулу АТФ.
Митохондрии содержат белки, РНК, тяжи ДНК, рибосомы, сходные с бактериальными, и различные растворенные вещества. ДНК существует в виде кольцевых молекул, располагающихся в одном или нескольких нуклеотидах.
Пластиды, наряду с вакуолями и клеточной оболочкой – характерные компоненты растительных клеток. Каждая пластида окружена собственной оболочкой, состоящей из двух элементарных мембран. Внутри пластид различают мембранную систему и более или менее гомогенное вещество – строму. Из пластид наибольшее значение имеют хлоропласты, в которых идет фотосинтез. Внутренняя структура хлоропласта довольно сложна. Строма пронизана развитой системой мембран, имеющих форму плоских пузырьков, называемых тилакоидами.
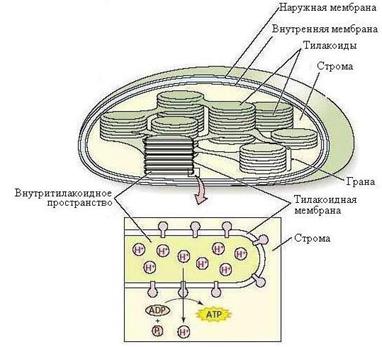
|
| Рис. 1.4. Схема строения хлоропласта |
Тилакоиды собраны в стопки – граны, напоминающие столбики монет имеющие в диаметре, как правило, 0,4–0,5 мкм (рис. 1.4). В одном хлоропласте имеется 10–50 таких гран.
Большая часть объема хлоропластов – строма. В строме содержатся ферменты, участвующие в фиксации углекислого газа и превращения его в различные продукты фотосинтеза.
Хлоропласты, в которых протекает фотосинтез, содержат хлорофиллы и каротиноиды. Размер – 4–5 мкм. В одной клетке мезофилла листа может содержаться 40–50 хлоропластов, в мм2 листа – около 500 000. В цитоплазме хлоропласты обычно располагаются параллельно клеточной оболочке.
Хлорофиллы и каротиноиды встроены в тилакоидные мембраны. Хлоропласты зеленых растений и водорослей часто содержат зерна крахмала и мелкие липидные (жировые) капли. Крахмальные зерна – это временные хранилища продуктов фотосинтеза. Они могут исчезнуть из хлоропластов, находящихся в темноте всего лишь 24 ч и появиться вновь уже через 3–4 ч после переноса растений на свет.
В изолированных хлоропластах осуществляется синтез РНК, который обычно контролируется только хромосомной ДНК. Образование хлоропластов и синтез находящихся в них пигментов в значительной степени контролируется хромосомной ДНК, малопонятным образом взаимодействующей с ДНК хлоропластов. Тем не менее, в отсутствие собственной ДНК хлоропласты не формируются.
Хлоропласты могут считаться основными клеточными органеллами, поскольку первыми стоят в цепи преобразования солнечной энергии, в результате которого мы получаем пищу и топливо. В хлоропластах протекают не только фотосинтез. Они участвуют в синтезе аминокислот и жирных кислот, служат хранилищем временных запасов крахмала.
Хромопласты (от греческого сhroma – цвет) – пигментированные пластиды. Многообразные по форме хромопласты не содержат хлорофилла, но синтезируют и накапливают каротиноиды, которые придают желтую, оранжевую и другую окраску. Корнеплоды моркови, плоды томатов окрашены пигментами, которые находятся в хромопластах.
Лейкопласты являются местом накопления запасного вещества – крахмала. Особенно много лейкопластов в клетках клубней картофеля. На свету лейкопласты могут преобразовываться в хлоропласты (клубни картофеля зеленеют). Осенью хлоропласты преобразуются в хромопласты и зеленые листья, и плоды желтеют и краснеют.
В отличие от пластид и митохондрий, которые ограничены двумя мембранами, микротельца представляют собой сферические органеллы, окруженные одной мембраной. Их диаметр колеблется от 0,5 до 1,5 мкм. Микротельца имеют гранулярное содержимое, иногда в них встречаются и кристаллические белковые включения. Микротельца обычно связаны с одним или двумя участками эндоплазматического ретикулума.
Некоторые микротельца, называемые пероксисомами, играют важную роль в метаболизме гликолевой кислоты, имеющей непосредственное отношение к фотодыханию. в зеленых листьях они связаны с митохондриями и хлоропластами. Другие микротельца, называемые глиоксисомами, содержат ферменты, необходимые для превращения жиров в углеводы, что происходит во многих семенах во время прорастания.
Микрофеламенты вместе с микротрубочками образуют гибкую сеть, называемую цитокселетом.
Микротрубочки – это тонкие цилиндрические структуры диаметром около 24 нм. Длина их варьирует. Каждая из микротрубочек состоит из субъединиц белка, называемого тубулином. Микротрубочки представляют собой динамические структуры, они регулярно разрушаются и образуются вновь на определенных стадиях клеточного цикла.
Функции их разнообразны. В растягивающихся и дифференцирующихся клетках микротрубочки, расположенные около внутренней поверхности плазматической мембраны, по-видимому, участвуют в образовании клеточной оболочки, контролируя упаковку целлюлозных микрофибрилл, которые откладываются цитоплазмой на растущую клеточную оболочку. Направление растяжения клетки определяется ориентацией целлюлозных микрофибрилл в оболочке. Микротрубочки направляют пузырьки диктиосом к формирующейся оболочке, подобно нитям веретена, которые образуются в делящейся клетке, и, вероятно, играют роль в формировании клеточной пластинки (первоначальной границы между дочерними клетками). Кроме того, микротрубочки – важный компонент жгутиков и ресничек, в движении которых, по-видимому, играют важную роль.
Микрофиламенты, подобно микротрубочкам, найдены практически во всех эукариотических клетках. Они представляют собой длинные нити толщиной 5–7 нм, состоящие из сократительного белка актина. Пучки микрофиламентов встречаются во многих клетках высших растений и играют определенную роль в движении цитоплазмы.
Все известные одноклеточные и многоклеточные организмы разделяются на две большие группы – прокаристы и эукариоты. К первым относятся бактерии и сине-зеленые водоросли (цианобактерии), ко вторым – зеленые растения, в том числе и остальные водоросли. Клетки прокариот (от греческого слова pro – до, karion – ядро) не имеют ядра. Другими словами можно сказать, что генетический материал (ДНК) прокариот находится в цитоплазме (отсутствует ядерная мембрана).
У эукариот (от греческого eu – настоящий, действительный, karion – ядро), наоборот, есть ядро, таким образом, у них генетический материал ограничен двойной мембраной (ядерной оболочкой) и образует отдельную клеточную структуру, которую легко узнать. Прокариоты отличаются от эукариот по целому ряду признаков.
Клетки эмбриональных тканей животных и растений в общем плане строения очень сходны, если не учитывать особенностей организации клеточной стенки у растений. Морфологические различия, связанные с образом жизни и способом питания, проявляются уже в дифференцированных клетках специализированных тканей растений и животных.
Вещества, составляющие пищу растений (углекислый газ, минеральные элементы и вода) и находятся вокруг растения в рассеянном виде. Зеленые растения на свету осуществляют автотрофный способ питания. Поэтому эволюционно сложились некоторые специфические особенности строения и роста растительных клеток. К ним относятся: 1) прочная полисахаридная клеточная стенка, окружающая клетку; 2) пластидная система (хлоропласты), возникшая в связи с указанным типом питания; 3) крупная центральная вакуоль, в зрелых клетках, играющая важную роль в поддержании тургорного давления клеток; 4) плазмодесмы – узкие каналы, обеспечивающие практически во всем растении непрерывность протоплазмы и возможность диффузии малых молекул из клетки в клетку; 5) наличие особого типа роста – роста растяжением. Кроме того, у делящихся растительных клеток нет центриолей.
Дата добавления: 2014-12-22; просмотров: 3202;