Мясо Хлеб Молоко
Рис.35.Кривые сокоотделения павловского желудочка на мясо, хлеб и молоко.
привести к повреждению слизистой оболочки желудка. Разрушению слизистого барьера и стимуляции секреции соляной кислоты способствует деятельность микроорганизмов Helicobacter pylori. В кислой среде и в условиях нарушенного слизистого барьера возможно переваривание элементов слизистой оболочки пепсином (пептический фактор язвообразования). Этому способствует также снижение секреции гидрокарбонатов и микроциркуляции крови в слизистой оболочке желудка.
Регуляция желудочной секреции. Вне пищеварения железы желудка выделяют небольшое количество желудочного сока. Прием пищи резко увеличивает его выделение. Это происходит за счет стимуляции желудочных желез нервными и гуморальными механизмами, составляющими единую систему регуляции. Стимулирующие и тормозные регуляторные факторы обеспечивают зависимость сокоотделения желудка от вида принимаемой пищи. Эта зависимость была впервые обнаружена в лаборатории И. П. Павлова в опытах на собаках с изолированным павловским желудочком, которым скармливалась различная пища. Объем и характер секреции во времени, кислотность и содержание в соке пепсинов определяются видом принятой пищи (рис.35).
Стимуляция секреции соляной кислоты обкладочными клетками осуществляется непосредственно и опосредованно через другие механизмы. Непосредственно стимулируют секрецию соляной кислоты обкладочными клетками холинергические волокна блуждающих нервов, медиатор которых - ацетилхолин (АХ) - возбуждает М-холинорецепторы базолатеральных мембран гландулоцитов. ЭффектыАХ и его аналогов блокируются атропином. Непрямая стимуляция клеток блуждающими нервами опосредуется также гастрином и гистамином.
Гастрин высвобождается из G-клеток, основное количество которых находится в слизистой оболочке пилорической части желудка. После хирургического удаления пилорической части желудочная секреция резко снижается. Высвобождение гастрина усиливается импульсами блуждающего нерва, а также местным механическим и химическим раздражением этой части желудка. Химическими стимуляторами G-клеток являются продукты переваривания белков - пептиды и некоторые аминокислоты, экстрактивные вещества мяса и овощей. Если рН в антральной части желудка понижается, что обусловлено повышением секреции соляной кислоты железами желудка, то высвобождение гастрина уменьшается, а при рН 1,0 прекращается и объем секреции резко понижается. Таким образом, гастрин принимает участие в саморегуляции желудочной секреции в зависимости от величины рН содержимого антрального отдела. Гастрин в наибольшей мере стимулирует париетальные гландулоциты желудочных желез и увеличивает выделение соляной кислоты.
К стимуляторам обкладочных клеток желудочных желез относится и гистамин, образующийся в ECL-клетках слизистой оболочки желудка. Высвобождение гистамина обеспечивается гастрином. Гистамин стимулирует гландулоциты, влияя на Н2-рецепторы их мембран и вызывая выделение большого количества сока высокой кислотности, но бедного пепсином.
Стимулирующие эффекты гастрина и гистамина зависят от сохранности иннервации желудочных желез блуждающими нервами: после хирургической и фармакологической ваготомии секреторные эффекты этих гуморальных стимуляторов понижаются. Желудочную секрецию возбуждают также всосавшиеся в кровь продукты переваривания белков.
Торможение секреции соляной кислоты вызывают секретин, ХЦК, глюкагон, ЖИП, ВИП, нейротензин, полипептид УУ, соматостатин, тиролиберин, энтерогастрон, АДГ, кальцитонин, окситоцин, простагландин ПГЕ2, бульбогастрон, кологастрон, серотонин (табл. ). Высвобождение некоторых из них в соответствующих эндокринных клетках слизистой оболочки кишечника контролируется свойствами химуса. В частности, торможение желудочной секреции жирной пищей в большой мере обусловлено влиянием на железы желудка ХЦК. Повышение кислотности содержимого двенадцатиперстной кишки тормозит выделение соляной кислоты железами желудка. Торможение секреции осуществляется рефлекторно, а также вследствие образования гормонов двенадцатиперстной кишки.
Механизм стимуляции и торможения секреции соляной кислоты различными нейротрансмиттерами и гормонами неодинаков. Так, АХ усиливает секрецию кислоты обкладочными клетками путем активации мембранной Nа+, К+-АТФазы, увеличения транспорта ионов Са2+ и эффектов повышенного внутриклеточного содержания цГМФ, высвобождения гастрина и потенцирования его влияния.
Гастрин усиливает секрецию соляной кислоты посредством гистамина, а также путем действия на мембранные рецепторы гастрина и усиления внутриклеточного транспорта ионов Са2+. Гистамин стимулирует секрецию обкладочных клеток черезих мембранные Н2-рецепторы и систему аденилатциклаза (АЦ) - цАМФ.
Стимуляторами секреции пепсиногена главными клетками являются холинергические волокна блуждающих нервов, гастрин, гистамин, симпатические волокна, оканчивающиеся на в-адренорецепторах, секретин и ХЦК. Усиление секреции пепсиногенов главными клетками желудочных желез осуществляется несколькими механизмами. Среди них увеличение переноса ионов Са2+ в клетку и стимуляция Nа+, К+-АТФазы; усиление внутриклеточного перемещения гранул зимогена, активация мембранной фосфорилазы, что усиливает их прохождение через апикальные мембраны, активация системы цГМФ и цАМФ.
Эти механизмы в неодинаковой мере активируются или тормозятся различными нейротрансмиттерами и гормонами, непосредственными и опосредованными влияниями их на главные клетки и секрецию пепсиногена. Показано, что гистамин и гастрин влияют, на него опосредованно - усиливают секрецию соляной кислоты, а снижение рН содержимого желудка через местный холинергический рефлекс усиливает секрецию главных клеток. Описано и прямое стимулирующее влияние на них гастрина. В высоких дозах гистамин тормозит их секрецию. ХЦК, секретин и в-адреномиметики непосредственно стимулируют секрецию главных клеток, но тормозят секрецию обкладочных, что свидетельствует о существовании на них разных рецепторов регуляторных пептидов.
Стимуляция секреции слизи мукоцитами осуществляется холинергическими волокнами блуждающих нервов. Гастрин и гистамин умеренно стимулируют мукоциты, видимо, в связи с удалением слизи с их мембран при выраженной секреции кислого желудочного сока. Ряд ингибиторов секреции соляной кислоты - серотонин, соматостатин, адреналин, дофамин, энкефалин, простагландин ПГЕ2 - усиливает секрецию слизи. Полагают, что ПГЕ2 усиливает секрецию слизи названными веществами.
При приёме пищи и пищеварении в усиленно секретирующих железах желудка кровоток возрастает, что обеспечивается действием холинергических нервных механизмов, пептидов пищеварительного тракта и местных вазодилататоров. В слизистой оболочке кровоток нарастает интенсивнее, чем в подслизистой основе и мышечном слое желудочной стенки.
Фазы желудочной секреции. Нервные, гуморальные факторы и паракринные механизмы тонко регулируют секрецию желез желудка, обеспечивают выделение определенного количества сока, кислото- и ферментовыделение в зависимости от количества и качества принятой пищи, эффективности ее переваривания в желудке и тонкой кишке. Происходящую при этом секрецию принято делить на три фазы.
Начальная секреция желудка возникает рефлекторно в ответ на раздражение дистантных рецепторов, возбуждаемых видом и запахом пищи, всей обстановкой, связанной с ее приемом (условнорефлекторные раздражения). Кроме того, секреция желудка возбуждается рефлекторно в ответ на раздражение принимаемой пищей рецепторов полости рта и глотки (безусловнорефлекторные раздражения). Эти рефлексы обеспечивают пусковые влияния на железы желудка. Желудочную секрецию, обусловленную этими сложными рефлекторными влияниями, принято называть первой, или мозговой, фазой секреции.
Механизмы первой фазы секреции желудка были изучены в опытах на эзофаготомированных собаках с фистулой желудка. При кормлении такой собаки пища выпадает из пищевода и не поступает в желудок, однако через 5-10 минут после начала мнимого кормления начинает выделяться желудочный сок. Аналогичные данные были получены при исследовании людей, страдающих сужением пищевода и подвергшихся вследствие этого операции наложения фистулы желудка. Жевание пищи вызывало у людей выделение желудочного сока.
Рефлекторные влияния на желудочные железы передаются через блуждающие нервы. После их перерезки у эзофаготомированной собаки ни мнимое кормление, ни вид и запах пищи не вызывают секреции. Если раздражать периферические концы перерезанных блуждающих нервов, то отмечается выделение желудочного сока с высоким содержанием в нем соляной кислоты и пепсина.
В стимуляцию желудочных желез в первую фазу включен и гастриновый механизм. Доказательством этого служит увеличение содержания гастрина в крови людей при мнимом кормлении. После удаления пилорической части желудка, где продуцируется гастрин, секреция в первую фазу понижается.
Секреция в мозговую фазу зависит от возбудимости пищевого центра и может легко тормозиться при раздражении различных внешних и внутренних рецепторов. Так, плохая сервировка стола, неопрятность места приема пищи снижают и тормозят желудочную секрецию. Оптимальные условия приема пищи положительно влияют на желудочную секрецию. Прием в начале еды сильных пищевых раздражителей повышает желудочную секрецию в первую фазу.
На секрецию первой фазы наслаивается секреция второй фазы, которая называется желудочной, так как обусловлена влиянием пищевого содержимого в период его нахождения в желудке. Наличие этой фазы секреции доказывается тем, что вкладывание пищи в желудок через фистулу, вливание через нее или зонд растворов в желудок, раздражение его механорецепторов вызывают отделение желудочного сока. Объем секреции при этом в 2-3 раза меньше, чем при естественном приеме пищи. Это подчеркивает большое значение пусковых рефлекторных влияний, осуществляемых преимущественно в первую фазу на желудочные железы. Во вторую фазу железы желудка испытывают в основном корригирующие влияния. Эти влияния путем усиления и ослабления деятельности желез обеспечивают соответствие секреции количеству и свойствам пищевого желудочного содержимого, т. е. осуществляют коррекцию секреторной деятельности желудка.
Сокоотделение при механическом раздражении желудка возбуждается рефлекторно с механорецепторов слизистой оболочки и мышечного слоя стенки желудка. Секреция резко уменьшается после перерезки блуждающих нервов. Кроме того, механическое раздражение желудка, особенно его пилорической части, приводит к высвобождению из G-клеток гастрина.
Повышение кислотности содержимого антральной части желудка тормозит высвобождение гастрина и снижает желудочную секрецию. В фундальной части желудка кислотность его содержимого рефлекторно усиливает секрецию, особенно выделение пепсиногена. Определенное значение в реализации желудочной фазы секреции имеет гистамин, значительное количество которого образуется в слизистой оболочке желудка.
Мясной бульон, капустный сок, продукты гидролиза белков при введении в тонкую кишку вызывают выделение желудочного сока. Нервные влияния с рецепторов кишечника на железы желудка обеспечивают секрецию в третью, кишечную, фазу. Возбуждающие и тормозные влияния из двенадцатиперстной и тощей кишки на железы желудка осуществляются с помощью нервных и гуморальных механизмов, корригирующих секрецию. Нервные влияния передаются с механо- и хеморецепторов кишечника. Стимуляция желудочных желез в кишечную фазу является прежде всего результатом поступления в двенадцатиперстную кишку недостаточно физически и химически обработанного содержимого желудка. В стимуляции желудочной секреции принимают участие всосавшиеся в кровь продукты гидролиза питательных веществ, особенно белков. Эти вещества могут возбуждать железы желудка опосредованно через гастрин и гистамин, а также непосредственно действуя на желудочные железы.
Торможение желудочной секреции в ее кишечную фазу вызывается рядом веществ в составе кишечного содержимого, которые по убывающей силе тормозного действия расположены в следующем порядке: продукты гидролиза жира, полипептиды, аминокислоты, продукты гидролиза крахмала, Н+ (рН ниже 3 оказывает сильное тормозное действие). Высвобождение в двенадцатиперстной кишке секретина и ХЦК под влиянием поступившего в кишечник содержимого желудка и образовавшихся продуктов гидролиза питательных веществ тормозит секрецию соляной кислоты, но усиливает секрецию пепсиногена. Желудочную секрецию тормозят и другие кишечные гормоны из группы гастронов и глюкагон, а также серотонин.
Влияние пищевых режимов на желудочную секрецию. В экспериментах на животных И. П. Павловым с сотрудниками, а затем И. П. Разенковым с сотрудниками показано, что секреция желудочных желез значительно изменяется в зависимости от характера питания. При длительном (30-40 дней) употреблении пищи, содержащей большое количество углеводов (хлеб, овощи), секреция уменьшается (в основном во вторую и третью фазы). Если животное длительный срок (30-60 дней) принимает пищу, богатую белками, например мясо, то секреция увеличивается, в особенности во вторую и третью фазы. При этом меняются не только объем и динамика во времени желудочной секреции, но и ферментативные свойства желудочного сока. А. М. Уголевым экспериментально установлено, что длительный прием растительной пищи повышает активность желудочного сока по отношению к белкам растительного происхождения («фитолитическая активность»), а преобладание в пищевом рационе животных белков повышает способность желудочного сока гидролизовать их («зоолитическая активность»). Это связано с изменением кислотности сока и соотношения в нем видов и свойств пепсинов.
Во время и в первые минуты после приема пищи желудок расслабляется - наступает пищевая рецептивная релаксация желудка, которая способствует депонированию пищи в желудке и его секреции (рис.36). Спустя некоторое время в зависимости от вида пищи сокращения усиливаются, при этом наименьшая сила сокращения отмечается в кардиальной части желудка и наибольшая - в антральной. Сокращения желудка начинаются на большой кривизне в непосредственной близости от пищевода, где находится кардиальный водитель ритма. Второй водитель ритма локализован в пилорической части желудка.
При баллонной гастрографии (рис.37) регистрируется три типа волн сокращений желудка: I - однофазные волны низкой амплитуды, давление колеблется от 1-2 до 5-10мм рт. ст., длительностью 5-20 с; II - однофазные волны большой амплитуды, давление составляет 40-80 мм рт. ст., длительностью 12- 60 с; III - сложные волны, возникают на фоне меняющегося исходного давления. Волны I и II типов носят перистальтический характер, поддерживают определенный тонус желудка, обеспечивают смешивание пищи с желудочным соком в непосредственной близости к слизистой оболочке желудка. Средняя частота этих волн 3 в 1 мин. В центральной части желудка содержимое не перемешивается, поэтому пища, принятая в разное время, располагается в желудке слоями (стратификация). Волны III типа характерны для пилорической части желудка, носят пропульсивный характер и участвуют в эвакуации содержимого в двенадцатиперстную кишку.
При регистрации внутрижелудочного давления методом открытых катетеров выявляются сокращения желудка двух типов: фазовые (тип А) и тонические (тип Б). Волны типа А быстрые, перистальтические, продолжительностью 10-20 с с частотой около 3 в 1 мин, волны типа Б - медленные, тонические, длительностью до 2 минут. Волны типа А бывают двух видов, первые имеют амплитуду и колебание давления от 1 до 15 мм рт. ст., у вторых амплитуда и колебание давления 16-30мм рт. ст. Тонические волны (тип Б) могут сочетаться и не сочетаться с фазовыми.
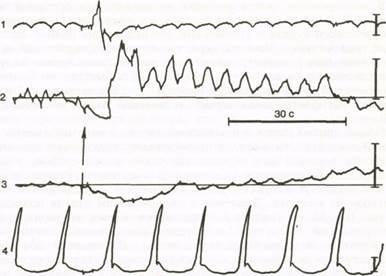
Рис. 36. Влияние глотания (указано стрелкой) на моторику пищевода (1), кар-диального сфинктера (2), фундального (3) и антрального (4) отделов желудка собаки. Высота вертикальных линий справа соответствует давлению 25 мм вод. ст.
Давление, мм рт.ст.
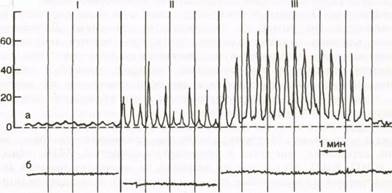
Рис. 37. Три типа волн сокращений желудка здорового человека (I-III).
а - сокращения желудка; б - пневмограмма.
В наполненном пищей желудке возникают три основных вида движений: перистальтические волны, систолические сокращения пилорического отдела и тонические, уменьшающие размер полости дна и тела желудка. Частота перистальтических сокращений около 3 в 1 мин; они распространяются от кардиальной части желудка к пилорической со скоростью около 1 см/с, быстрее по большой, чем по малой кривизне, длятся около 1,5 с. В пилорической части скорость распространения перистальтической волны увеличивается до 3-4 см/с.
После приема пищи и в зависимости от ее вида параметры моторной деятельности желудка приобретают характерную динамику. В течение первого часа перистальтические волны слабые, в дальнейшем они усиливаются (в пилорическом отделе увеличиваются их амплитуда и скорость распространения), проталкивая пищу к выходу из желудка. Давление в пилорическом отделе повышается до 10-25 см вод.ст., открывается сфинктер привратника (пилорический сфинктер), и порция желудочного содержимого переходит в двенадцатиперстную кишку. Оставшееся (большее) количество его возвращается в проксимальную часть пилорического отдела желудка. Такие движения желудка обеспечивают перемешивание и перетирание (фрикционный эффект) пищевого содержимого, его гомогенизацию. Характер, интенсивность, временная динамика моторики зависят от количества и вида пищи, от эффективности ее переваривания в желудке и кишечнике, обеспечивается регуляторными механизмами.
Рассмотрим регуляцию моторики желудка. Раздражение блуждающих нервов и выделение АХ усиливают моторику желудка: увеличивают ритм и силу сокращений, ускоряют движение перистальтических волн. Влияния блуждающих нервов могут оказывать и тормозной эффект: рецептивная релаксация желудка, снижение тонуса пилорического сфинктера. Раздражение симпатических нервов и активация б-адренорецепторов тормозят моторику желудка: уменьшают ритм и силу его сокращений, скорость движения перистальтической волны. Описаны и стимулирующие б- и в-адренорецепторные влияния (например, на пилорический сфинктер). Двунаправленные влияния осуществляются пептидергическими нейронами. Названные типы влияний осуществляются рефлекторно при раздражении рецепторов рта, пищевода, желудка, тонкой и толстой кишки. Замыкание рефлекторных дуг осуществляется на различных уровнях ЦНС, в периферических симпатических ганглиях и интрамуральной нервной системе.
В регуляции моторики желудка велико значение гастроинтестинальных гормонов. Моторику желудка усиливают гастрин, мотилин, серотонин, инсулин, а тормозят - секретин, ХЦК, глюкагон, ЖИП, ВИП. Механизм их влияний на моторику прямой (непосредственно на мышечные пучки и миоциты) и опосредованный через интрамуральные нейроны. Моторика желудка зависит от уровня его кровоснабжения и сама влияет на него, изменяя сопротивление кровотоку при сокращениях желудка.
Дата добавления: 2014-12-22; просмотров: 1769;