КАТАБОЛИЗМ АМИНОКИСЛОТ.
80% аминокислот, которые поступают в организм из желудочно-кишечного тракта, используются для синтеза белков. Остальные 20% вступают в метаболические процессы. Все эти процессы можно разделить на 2 группы:
1. Общие пути катаболизма аминокислот (для всех аминокислот они одинаковы). В них принимает участие общая часть молекулы аминокислоты.
2. Специфические пути метаболизма для каждой отдельной аминокислоты (разные для разных аминокислот) - участвуют радикалы аминокислот. Это - особенности обмена отдельных аминокислот.
ОБЩИЕ ПУТИ КАТАБОЛИЗМА АМИНОКИСЛОТ
1. Декарбоксилирование
2. Дезаминирование
3. Трансаминирование (переаминирование)
ДЕКАРБОКСИЛИРОВАНИЕ
В природе встречаются разные типы декарбоксилирования аминокислот. В организме человека происходит только окислительное декарбоксилирование. Ферменты - декарбоксилазы. Их простетическая группа представлена пиридоксальфосфатом - это активная форма витамина В6:
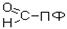
|
В реакциях декарбоксилирования участвует альдегидная группа пиридоксальфосфата:
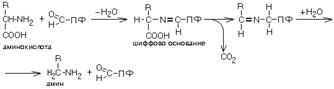 Аминокислота соединяется с активным центром фермента, в состав которого входит альдегидная группа ПФ. Образуются Шиффовы основания (альдимины и кетимины). В результате СООН-группа становится лабильной и отщепляется в виде СО2. Далее происходит гидролиз до соответствующего амина. Эта реакция необратима. Отнятие СО2 происходит без окисления.
Аминокислота соединяется с активным центром фермента, в состав которого входит альдегидная группа ПФ. Образуются Шиффовы основания (альдимины и кетимины). В результате СООН-группа становится лабильной и отщепляется в виде СО2. Далее происходит гидролиз до соответствующего амина. Эта реакция необратима. Отнятие СО2 происходит без окисления.
Субстратная специфичность декарбоксилаз очень разная.
|
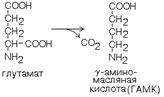
1. ГЛУТАМАТДЕКАРБОКСИЛАЗА - высокоспецифичный фермент. Работает в клетках серого вещества головного мозга. Катализирует реакцию превращения глутаминовой кислоты в гамма-аминомасляную кислоту (ГАМК).
ГАМК является медиатором тормозных импульсов в нервной системе. ГАМК и ее аналоги применяются в медицине как нейротропные средства для лечения эпилепсии и других заболеваний.
2. ОРНИТИН-ДЕКАРБОКСИЛАЗА - высокоспецифичный фермент. Катализирует превращение орнитина в путресцин:
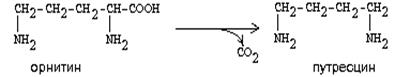
|
Образующийся ПУТРЕСЦИН (диаминобутан) является трупным ядом. В результате присоединения остатков пропиламина из путресцина могут образоваться СПЕРМИН и СПЕРМИДИН, содержащие 3 (у спермина) или 4 (у спермидина) имино- или аминогруппы.

|
Спермин и спермидин относятся к группе биогенных полиаминов. Введение полиаминов в организм снижает температуру тела и кровяное давление. Полиамины принимают участие в процессах пролиферации клеток и роста тканей, а также в регуляции биосинтеза белка. Они являются ингибиторами некоторых ферментов, в том числе протеинкиназ.
Орнитиндекарбоксилаза - это первый фермент на пути образования путресцина и остальных полиаминов, это регуляторный фермент процесса.
В культуре клеток добавление некоторых гормонов ускоряет биосинтез орнитиндекарбоксилазы в 10-200 раз.
Период полужизни орнитиндекарбоксилазы - 10 минут.
Добавление в культуру клеток самих полиаминов приводит к индукции биосинтеза другого белка - ингибитора орнитиндекарбоксилазы. При раковых заболеваниях обнаружено резкое увеличение секреции полиаминов и повышение их экскреции с мочой.
3. ГИСТИДИНДЕКАРБОКСИЛАЗА
Этот фермент имеет абсолютную субстратную специфичность - превращает гистидин в гистамин:
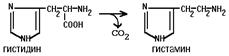
|
Гистамин является медиатором и содержится в нервных клетках и в тучных клетках. Обладают сильным сосудорасширяющим действием. Особенно много его выделяется в очаге воспаления. Гистамин играет важную роль в проявлении аллергических реакций.
Известно 2 типа рецепторов к гистамину:H1 и H2.
Эффекты гистамина:
- расширение капилляров и повышение сосудистой проницаемости;
- понижение артериального давления;
- повышение тонуса (спазм) гладких мышц - в том числе гладкой мускулатуры бронхов;
- усиление секреции желудочного сока;
Некоторые из этих эффектов позволяют гистамину принимать участие в формировании аллергических проявлений.
Антигистаминные препараты применяются с целью предотвратить образование гистамина и обладают противовоспалительным и антиаллергическим действием. По механизму действия некоторые из них являются ингибиторами гистидин-декарбоксилазы, а другие конкурируют с гистамином за взаимодействие с рецепторами клеток.
Например, лекарственный препарат циметидин и его аналоги блокируют Н2-рецепторы и таким образом понижают секрецию желудочного сока. Применяются при лечении язвенной болезни желудка.
Блокаторы Н1-рецепторов используются в основном как противоаллергические средства - димедрол, тавегил, супрастин, пипольфен, грандаксин. Некоторые из этих препаратов вызывают сонливость.
4. ДЕКАРБОКСИЛАЗА АРОМАТИЧЕСКИХ АМИНОКИСЛОТ
Имеет широкую субстратную специфичность. Превращает несколько разных аминокислот:
а) триптофан - в триптамин
б) 5-окситриптофан - в триптамин (серотонин)
в) 3,4-диоксифенилаланин - в дофамин
г) гистидин - в гистамин
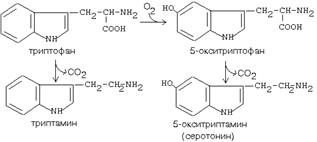
Серотонин вырабатывается в нервной ткани. Некоторые виды головных болей (мигрени) связаны с избыточной выработкой серотонина. Серотонин сужает сосуды, регулирует свертывание крови. Обладает антиаллергическим действием. Триптамин обладает сходным эффектом.
Аминокислота фенилаланин может в результате окисления присоединять две ОН-группы в кольце и превращаться в диоксифенилаланин (ДОФА). Из него под действием ДЕКАРБОКСИЛАЗЫ АРОМАТИЧЕСКИХ АМИНОКИСЛОТ образуется дофамин. Дофамин является предшественником катехоламинов - норадреналина и адреналина.
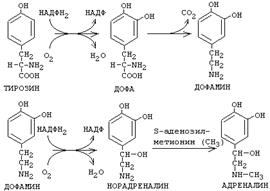
|
Кроме функции предшественника, ДОФАмин имеет свои специфические функции. Если ДОФА метилируется, то образуется a-метил-ДОФА. Это соединение является сильным ингибитором декарбоксилазы ароматических аминокислот. Применяется как лекарственный препарат для понижения артериального давления (называется - альдомет).
БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ РЕАКЦИЙ ДЕКАРБОКСИЛИРОВАНИЯ АМИНОКИСЛОТ
1. Реакции необратимы - приводят к необратимому распаду аминокислот.
2. Образуется значительное количество СО2 - конечного продукта метаболизма, который выводится из организма.
3. Образуются амины, которые обладают высокой биологической активностью. Поэтому такие амины называют биологически активными или биогенными аминами. Они являются медиаторами, с помощью которых сигнал передается от одной клетки к другой и от одной молекулы к другой.
ИНАКТИВАЦИЯ БИОГЕННЫХ АМИНОВ
Если биогенные амины обладают высокой биологической активностью, то они должны быстро разрушаться после выполнения своей функции.
В организме имеются механизмы, позволяющие разрушать биогенные амины.
МЕХАНИЗМЫ ИНАКТИВАЦИИ:
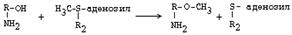
|
1. Метилирование по оксигруппам тех аминов, которые такие группы содержат, либо включают оксигруппы в свою молекулу после гидроксилирования.
Ферменты - О-МЕТИЛТРАНСФЕРАЗЫ. Они переносят метильную группу на кислород. Источник метильного радикала: S-Аденозилметионин.
После присоединения аденильного остатка АДФ к сере метионина, метильная группа метионина становится очень мобильной и легко переносится на разные вещества. В том числе и на кислород оксигрупп.
2. Окисление амина по аминогруппе с целью дезаминирования.
Главный путь инактивации биогенных аминов - их окисление под действием оксидаз с отщеплением аминогруппы. В результате исчезает биологическая активность амина.

Оксидазы биогенных аминов: моноаминооксидаза (МАО), диаминооксидаза (ДАО), полиаминооксидаза.
Оксидазы отнимают два протона и два электрона и передают их сразу на кислород. Образуется перекись водорода, а амин превращается в ИМИН. Этот имин легко гидролизуется без участия фермента и превращается в альдегид. Простетической группой ферментов оксидаз является ФАД или ФМН, т.е. они являются флавопротеинами.
Вторая реакция (гидролиз) необратима. Образовавшийся в итоге альдегид легко окисляется до карбоновой кислоты, которая распадается до СО2 и H2O. МАО в клетке больше, чем ДАО.
Угнетение МАО приволит к замедлению распада биогенных аминов. Такие лекарства продлевают период существования биогенных аминов, что особенно важно при их недостатке.
Эти вещества играют роль антидепрессантов и используются, в частности, при лечении шизофрении.
Аминокислоты декарбоксилируются в цитоплазме, а окисление аминов происходит в наружной мембране митохондрий. Поскольку реакция декарбоксилирования аминокислот и разрушение биогенных аминов происходят не одновременно, то биогенные амины могут некоторое время существовать и выполнять свою биологическую функцию.
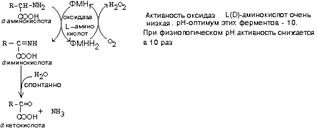
ДЕЗАМИНИРОВАНИЕ АМИНОКИСЛОТ
У человека происходит в основном путем окислительного дезаминирования. Эти реакции протекают с помощью двух ферментов:
- оксидаза Д-аминокислот
- оксидаза L-аминокислот
|
Эти ферменты обладают групповой стереоспецифичностью. Оксидазы отнимают протоны и электроны от аминокислот с помощью такого же механизма, как и оксидазы, обеспечивающие дезаминирование биогенных аминов. Эти ферменты являются флавопротеинами и содержат в качестве простетической группы ФАД или ФМН:
На первой стадии образуется иминокислота, а затем, после спонтанного гидролиза образуется альфа-кетокислота.
Кроме оксидаз имеется еще один фермент, катализирующий окислительное дезаминирование глутаминовой кислоты - глутамат-дегидрогеназа (глутаматДГ).
Этот фермент является НАД-зависимым и обладает высокой активностью (как и другие НАД-зависимые дегидрогеназы). В этом его отличие от оксидаз аминокислот, которые медленно превращают аминокислоты в физиологических условиях (поэтому в клетке сохраняется большинство аминокислот). Так как глутамат-ДГ является никотинамидной, то отнимаемые протоны и электроны не передаются сразу на кислород, а транспортируются по полной цепи МтО с образованием воды и параллельным образованием трех молекул АТФ.
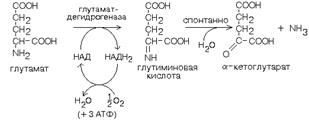 Глутамат-ДГ обладает высокой активностью и этим отличается от МАО и ДАО. Глутамат-ДГ является регуляторным ферментом - он ингибируется избытком АТФ, и активируется избытком АДФ.
Глутамат-ДГ обладает высокой активностью и этим отличается от МАО и ДАО. Глутамат-ДГ является регуляторным ферментом - он ингибируется избытком АТФ, и активируется избытком АДФ.
БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ РЕАКЦИЙ ДЕЗАМИНИРОВАНИЯ
1. Реакции дезаминирования необратимы, как и реакции декарбоксилирования - дезаминирование тоже может играть роль первого этапа на путях распада аминокислот.
2. Один из непосредственных продуктов дезаминирования -конечный продукт метаболизма аммиак. Это токсическое вещество. Поэтому клетки должны затрачивать энергию, чтобы обезвредить аммиак до безвредных продуктов, которые выводятся из организма.
3. Другой продукт реакции дезаминирования - альфа-кетокислота.
Все образующиеся альфа-кетокислоты легко расщепляются дальше до СО2 и Н2О (например, аланин превращается в ПВК (путем дезаминирования; аспартат - в ЩУК; глутаминовая кислота - в альфа-кетоглутаровую). Большинство альфа-кетокислот тем или иным путем превращаются в кислоты, которые являются промежуточными метаболитами ЦТК:
- в альфа-кетоглутаровую;
- в янтарную;
- фумаровую;
- щавелево-уксусную. Все эти метаболиты могут в организме трансформироваться в углеводы, перед этим превращаясь в ПВК. Поэтому большинство аминокислот относится к группе, которая называется ГЛЮКОГЕННЫМИ АМИНОКИСЛОТАМИ (их 17). Только 3 аминокислоты не могут превращаться в ПВК, но превращаются в Ац-КоА - КЕТОГЕННЫЕ АМИНОКИСЛОТЫ: лейцин, лизин, триптофан). Они могут прямо трансформироваться в жирные кислоты или в кетоновые тела.
 Метаболические пути, в которые вступают аминокслоты после дезаминирования, уже не являются собственно путями метаболизма аминокислот, а являются универсальными и для аминокислот, и для углеводов, и для жиров.
Метаболические пути, в которые вступают аминокслоты после дезаминирования, уже не являются собственно путями метаболизма аминокислот, а являются универсальными и для аминокислот, и для углеводов, и для жиров.
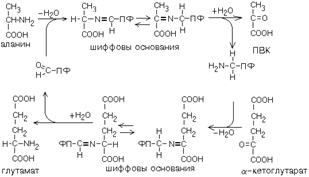
ТРАНСАМИНИРОВАНИЕ
Эта реакция заключается в том, что аминокислота и кетокислота обмениваются друг с другом своими функциональными группами при альфа-углеродном атоме. В результате вступившая в реакцию аминокислота превращается в соответствующую альфа-кетокислоту, а кетокислота становится аминокислотой.
Эту реакцию катализируют ферменты под названием ТРАНСАМИНАЗЫ (АМИНОТРАНСФЕРАЗЫ). Коферментом всех трансаминаз является активная форма витамина В6 - пиридоксальфосфат (фосфопиридоксаль).
ГЛАВНЫЕ ОСОБЕННОСТИ РЕАКЦИИ ТРАНСАМИНИРОВАНИЯ:
1. Это циклический процесс, все стадии которого катализируются одним и тем же ферментом - трансаминазой данной пары кислот. В этот цикл вступает одна аминокислота и кетокислота (в приведенном примере - это глутамат и ПВК). Образуются другая альфа-кетокислота и аминокислота (в приведенном примере - альфа-кетоглутарат и аланин).
2. Все стадии этого процесса обратимы. Поэтому весь цикл в целом может протекать как в прямом, так и в обратном направлении. Направление этого цикла зависит от соотношения концентраций участников реакции - всех четырех кислот. Резко повысив концентрацию какой-либо одной из кислот, можно направить реакцию по определенному направлению.
3. Каждая трансаминаза обычно специфична для одной пары субстратов и соответствующей ей пары продуктов и все стадии реакции катализируются только одним ферментом. По участникам процесса для данного фермента дают ему название:
Фермент, который катализирует приведенную реакцию, можно назвать: Аланинаминотрансфераза (АлТ) (глутаминово-пировиноградная трансаминаза (ГПТ)).
4. Трансаминазы обычно обладают высокой субстратной специфичностью и высокой активностью. Наиболее активными в клетках являются те трансаминазы, для которых хотя бы один из субстратов является дикарбоновой альфа-кетокислотой - это АлТ - аланинаминотрансфераза (ГПТ) и АсТ - аспартатаминотрансфераза (ГЩТ).
Определение активности этих ферментов в сыворотке крови имеет важное значение в ДИФФЕРЕНЦИАЛЬНОЙ ДИАГНОСТИКЕ таких заболеваний, как ВИРУСНЫЙ ГЕПАТИТ и ИНФАРКТ МИОКАРДА.
АсТ и АлТ являются внутриклеточными ферментами. Поэтому в норме их активность в крови очень мала. При вирусном гепатите или при инфаркте миокарда наблюдается разрушение клеток печени или миокарда соответственно. Поэтому в крови будет наблюдаться повышение активности и АлТ, и АсТ, но неодинаковое для каждого из ферментов.
В клетках сердечной мышцы (миокарда) содержится гораздо больше АсТ, чем АлТ, а в клетках печени - наоборот: АлТ намного больше, чем АсТ. Поэтому ПРИ ИНФАРКТЕ МИОКАРДА АКТИВНОСТЬ АСТ в крови БУДЕТ ЗНАЧИТЕЛЬНО ВЫШЕ, ЧЕМ АЛТ, А ПРИ ВИРУСНОМ ГЕПАТИТЕ АКТИВНОСТЬ АЛТ БУДЕТ ВЫШЕ, ЧЕМ АСТ.
Поэтому определение активности этих ферментов в крови помогает врачам в постановке правильного диагноза.
БИОЛОГИЧЕСКАЯ РОЛЬ РЕАКЦИЙ ТРАНСАМИНИРОВАНИЯ.
1. ОБЕСПЕЧИВАЮТ СИНТЕЗ НОВЫХ АМИНОКИСЛОТ ИЗ ЧИСЛА ЗАМЕНИМЫХ. Из заменимых аминокислот также могут образоваться необходимые клетке кетокислоты.
Эта функция позволяет ругулировать содержание различных аминокислот в клетках организма (корректировка аминокислотного состава клеток).
В основе этой роли - полная обратимость реакции трансаминирования. Для характеристики направления каждой конкретной реакции трансаминирования и определения положения равновесия данной реакции вычисляют ее КОНСТАНТУ РАВНОВЕСИЯ (Кравновесия):
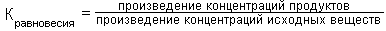
Для большинства реакций трансаминирования эта константа равновесия близка к единице.
 При избытке какой-либо из кислот соотношение быстро восстанавливается трансаминазой за счет другой пары. Недостающее количество какой-либо из кислот может быть взято клеткой из других метаболических путей (например, альфа-кетоглутарат может быть взят из ЦТК). Избыток какой-либо из кислот может быть ликвидирован другими ферментами (например, избыточное количество глутамата окисляется глутаматдегидрогеназой).
При избытке какой-либо из кислот соотношение быстро восстанавливается трансаминазой за счет другой пары. Недостающее количество какой-либо из кислот может быть взято клеткой из других метаболических путей (например, альфа-кетоглутарат может быть взят из ЦТК). Избыток какой-либо из кислот может быть ликвидирован другими ферментами (например, избыточное количество глутамата окисляется глутаматдегидрогеназой).
2. ОБЕСПЕЧИВАЮТ ПРОТЕКАНИЕ РЕАКЦИЙ КОСВЕННОГО ДЕЗАМИНИРОВАНИЯ.
3. ОБЕСПЕЧИВАЮТ СИНТЕЗ МОЧЕВИНЫ
КОСВЕННОЕ ДЕЗАМИНИРОВАНИЕ (ТРАНСДЕЗАМИНИРОВАНИЕ).
Оксидазы аминокислот в клетках организма человека имеют очень низкую активность. Поэтому прямое окислительное дезаминирование аминокислот почти не протекает. Исключением является никотинамидный фермент глутаматдегидрогеназа, который легко дезаминирует глутаминовую кислоту, расщепляя ее до NH3 и альфа-кетоглутарата. Альфа-кетоглутарат может легко вступать в реакции трансаминирования с другими аминокислотами, превращаясь обратно в глутаминовую кислоту.
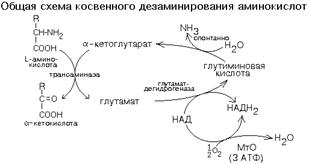 Значит, если аминокислота не может дезаминироваться прямо, то она может дезаминироваться косвенно с участием пары "альфа-кетоглутарат/глутамат".
Значит, если аминокислота не может дезаминироваться прямо, то она может дезаминироваться косвенно с участием пары "альфа-кетоглутарат/глутамат".
Некоторые аминокислоты не имеют "своей собственной" трансаминазы, которая могла бы перенести их аминогруппу на альфа-кетоглутарат (лизин, треонин, пролин, оксипролин), но для этих аминокислот есть специальные реакции, в результате которых аминогруппа этих аминокислот оказывается в составе глутаминовой кислоты.
Таким образом, глутаминовая кислота работает в паре с альфа-кетоглутаровой: альфа-кетоглутарат принимает у разных аминокислот азот в форме аминогруппы, превращаясь в глутамат; глутамат легко дезаминируется прямым путем высокоактивной глутаматдегидрогеназой с образованием аммиака.
Общий итог косвенного дезаминирования такой же, как и результат прямого окислительного дезаминирования.
Это вариант дезаминирования, который протекает в две стадии:
а) трансаминирование с участием альфа-кетоглутаровой кислоты;
б) дезаминирование образовавшейся на первой стадии глутаминовой кислоты.
Косвенное дезаминирование является наиболее важным, т.к. именно таким путем дезаминируются большинство аминокислот живого организма. Это обусловлено тем, что фермент глутамат-ДГ более активен, чем оксидазы аминокислот.
ПРЕИМУЩЕСТВА КОСВЕННОГО ДЕЗАМИНИРОВАНИЯ ПЕРЕД ПРЯМЫМ:
1. В этом процессе не образуется Н2О2, в отличие от прямого дезаминирования, а образуется Н2О и 3 молекулы АТФ (энергия).
2. Косвенное дезаминирование обеспечивает гораздо более быстрое дезаминирование (благодаря высокой активности фермента глутамат-ДГ и трансаминазы);
3. Косвенное дезаминирование протекает не только быстрее, но скорость этого процесса может регулироваться. Возможность регуляции косвенного дезаминирования обусловлена обратимостью процесса и автономной саморегуляцией. Аллостерическими регуляторами глутаматдегидрогеназы являются АДФ (сильный активатор) и АТФ (ингибитор). Ингибирующее действие на этот фермент оказывают также ГТФ и НАДН2.
Если АТФ начинает расходоваться больше, чем образуется в процессе биоокисления в митохондриях, то после активации глутамат-ДГ происходит смещение реакций в сторону образования АТФ, благодаря тому, что глутамат начинает быстрее дезаминироваться, а образующиеся кетокислоты, либо сразу, либо после определенных превращений, могут также вступать на путь митохондриального окисления. Это наблюдается при недостатке жирных кислот или углеводов. То есть при недостатке жиров и углеводов в пище, в качестве дополнительного источника энергии организм начинает использовать аминокислоты. Нередко приходится при голодании разрушать собственные белки. Т.е. при похудании в организме разрушаются не только жиры и углеводы, но и собственные белки. Присутствие достаточного количества жиров и углеводов в пище позволяет сберечь белки от нерационального использования.
СИНТЕЗ ЗАМЕНИМЫХ АМИНОКИСЛОТ В ОРГАНИЗМЕ
Иногда в организме возникает ситуация, когда потребность в тех или иных аминокислотах больше, чем их поступление с пищей. Допустим, организму требуется 7 граммов аланина и 15 граммов глутамата, а с пищей поступило всего лишь 5 граммов аланина, но зато 20 граммов глутамата. Тогда организм, используя механизм трансаминирования, может синтезировать недостающее количество аланина из ПВК, который синтезируется в бих количествах (сотни граммов в сутки). Таким образом, в организме постоянно происходит корректировка аминокислотного состава, но это не уменьшает потребности организма в пищевом белковом азоте. Эта корректировка осуществляется с использование соответствующих альфа-кетокислот, образующихся в ходе метаболизма углеводов и жиров.
Незаменимые аминокислоты не могут образуются в организме, потому что в ходе распада углеводов и жиров не образуется соответствующих им альфа-кетокислот.
Подводя итог, можно сказать, что в результате общих путей катаболизма аминокислот, они разрушаются до СО2, NH3 и безазотистых фрагментов, которые также могут разрушаться до СО2 и Н2О.
Аммиак подвергается реакциям обезвреживания.
ОБЕЗВРЕЖИВАНИЕ АММИАКА. СИНТЕЗ МОЧЕВИНЫ (ОРНИТИНОВЫЙ ЦИКЛ).
Только у рыб аммиак является конечным продуктом распада и выводится из организма. У птиц и рептилий, конечный продукт катаболизма азотсодержащих соединений - мочевая кислота, а не аммиак. У млекопитающих образующийся аммиак превращается в мочевину - это полный амид угольной кислоты:
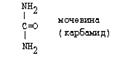
|
Образование мочевины происходит в печени в результате орнитинового цикла (открыт Г.Кребсом).
1. В матриксе митохондрий соединяются СО2 и NH3 (при этом расходуются две молекулы АТФ).
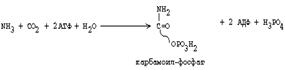
|
Синтез мочевины представляет собой циклический процесс, в который вступают предварительно синтезированный карбамоил-фосфат и аспартат, а образуются фумарат и мочевина.
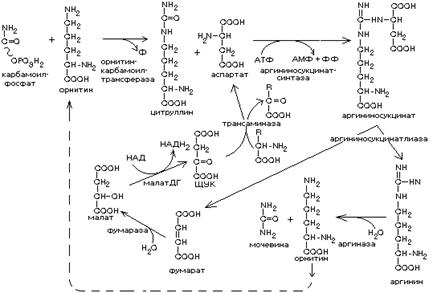
Мочевина синтезируется из одной молекулы СО2, одной молекулы NH3 и аминогруппы аспартата. Из фумарата в реакциях ЦТК вновь образуется щавелевоуксусная кислота, которая может вступать в трансаминирование с другими аминокислотами и превращаться в аспартат.
Таким образом, в орнитиновом цикле существуют два сопряженных цикла:
а) образование мочевины; б) регенерация аспартата.
В добавление к лекции по общим путям обмена аминокислот можно сказать, что еще одна, третья по счету функция трансаминирования - это перенос аминогруппы с аминокислот для синтеза мочевины без промежуточного выделения аммиака.
При синтезе мочевины расходуется в сумме 4 молекулы АТФ. Мочевина - это нетоксичное вещество, которое легко выводится из организма с мочой. Накопление мочевины в крови выше нормы происходит только при нарушениях функции почек.
Синтез мочевины происходит только в печени, а аммиак образуется в разных тканях. Значит, должен быть специальный механизм транспорта аммиака в безвредной для организма форме: это МЕХАНИЗМ ВРЕМЕННОГО ОБЕЗВРЕЖИВАНИЯ АММИАКА.
Обеспечивается ферментом глутамин-синтетазой, которая присоединяет с затратой АТФ дополнительную аминогруппу к гамма-карбоксигруппе:
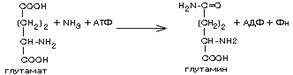
|
Аминогруппа может также присоединяться и к аспартату - к бета-карбоксигруппе.
Особенно важны реакции временного обезвреживания аммиака в нервной ткани (очень чувствительной к токсическому влиянию аммиака). Со значительной скоростью протекают они и в мышечной ткани.
Образующиеся амиды переносятся в печень, где отдают амидный азот на синтез мочевины. В почках некоторое количество аммиака из амидов может выделяться в свободном виде. Обычно такого аммиака образуется немного, но если в организме - ацидоз - сдвиг рН в кислую сторону, то этот аммиак начинает выделяться с мочой в больших количествах. Аммиак позволяет частично нейтрализовать кислоты, которые из крови при ацидозе попадают в мочу. Таким образом, глутамин и аспарагин являются транспортными формами аммиака и в составе своей молекулы переносят его из разных тканей к печени и к почкам.
СПЕЦИФИЧЕСКИЕ ПУТИ ОБМЕНА ОТДЕЛЬНЫХ АМИНОКИСЛОТ.
Эти пути обмена определяются различиями в строении радикалов аминокислот /АК/, поэтому они разнообразны и многочисленны. Вступая в эти специфические реакции, АК-ты принимают участие во многих важных процессах:
а) в синтезе гормонов и нейромедиаторов,
б) в синтезе простетических групп сложных белков - хромопротеинов и нуклеопротеинов,
в) в синтезе сложных липидов,
г) синтезе веществ, содержащих макроэргическую связь и являющихся источником энергии для клеток,
д) в обезвреживании токсических веществ.
Вступая в эти процессы, АК-ты участвуют в них как всей своей молекулой, так и своими отдельными фрагментами (или группировками). Ими являются аминогруппа (NH2) и одноуглеродные фрагменты: -СН3, -CH2-, -С=О, -СН= , -СН2ОН.
Одноуглеродные фрагменты образуются из простейших аминокислот - глицина, серина, аланина и треонина под действием ферментов, коферментом которых является тетрагидрофолиевая кислота/ТГФК/. ТГФК - производное витамина Вс (фолиевой кислоты), она образуется из фолиевой кислоты в результате ее восстановления с помощью НАДФН2. Одноуглеродный фрагмент, соединенный с ТГФК называют активным одноуглеродным фрагментом или " активный С1".
ОБМЕН ГЛИЦИНА И СЕРИНА.
Это заменимые аинокислоты, которые превращаются друг в друга.
Основным путем распада глицина является его распад на СО2, Н2О и метилен-ТГФК (активный С1):
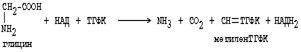
|
Из метилен-ТГФК могут образовываться все другие формы активного С1: формил-ТГФК, метил-ТГФК, метен-ТГФК, оксиметил-ТГФК в результате реакций окисления или восстановления метилен-ТГФК.
Активный С1, образовавшийся из глицина участвует в синтезе пуриновых азотистых оснований. Кроме того, глицин всей своей молекулой участвует в синтезе гема гемоглобина и других гемопротеинов, в синтезе пуриновых азотистых оснований, в синтезе парных желчных кислот (гликохолевая кислота), в синтезе креатина, в синтезе трипептида глютатиона. Также глицин в печени участвует в обезвреживании бензойной кислоты, которая превращается в гиппуровую кислоту: 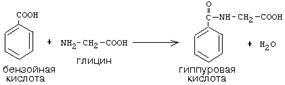
Основным путем распада серина является его превращение в метилен-ТГФК и глицин:
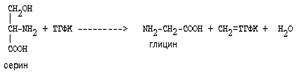
|
Всей своей молекулой серин участвует в синтезе сложных липидов - фосфолипидов. Он участвует в синтезе фосфатидилсерина, который декарбоксилируется и превращается в фосфатидилэтаноламин, который после метилирования превращается в фосфатидилхолин. В реакции метилирования участвует активная форма метионина - S-аденозилметионин.
Серин в составе белков-ферментов участвует в формировании каталитического центра фермента, например, в сериновых протеазах: трипсине, химотрипсине и др. Также серин участвует в формировании гидратной оболочки белков, потому что является полярной аминокислотой.
ОБМЕН СЕРУСОДЕРЖАЩИХ АМИНОКИСЛОТ: МЕТИОНИНА И ЦИСТЕИНА.
Метионин - это незаменимая аминокислота, а цистеин - заменимая.
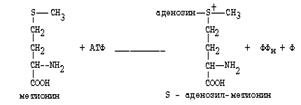
|
Главной особенностью обмена метионина является то, что из него тоже образуется активный С1 в виде СН3-группы, которая участвует в различных синтезах. Однако, этот активный С1 образуется без участия ТГФК. Чтобы стать источником СН3- группы, метионин подвергается активации с участием АТФ. В результате этой реакции от АТФ отщепляются все три остатка фосфорной кислоты, а аденозин присоединяется к атому серы метионина. Так образуется активная форма метионина - S- аденозил-метионин.
S-аденозил-метионин участвует в реакциях трансметилирования. Наиболее важный из них синтез фосфатидилхолина из фосфатидилэтаноламина, обезвреживание биогенных аминов с участием О-метилтрансфераз, синтез адреналина из норадреналина, синтез ацетилхолина из холина и Ацетил-КоА, синтез креатина, который в виде креатинфосфата является резервной формой макроэргических связей и участвует в обеспечении нервной ткани и работающей мышцы АТФ.
Синтез креатина.
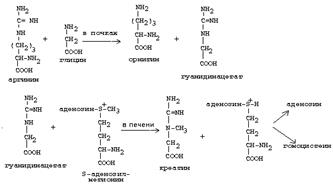
|
В синтезе креатина участвуют и другие аминокислоты - аргинин и глицин. В почках из аргинина и глицина образуется гуанидинацетат, который метилируется в печени с участием S-аденозил-метионина и в результате образуется креатин:
Гомоцистеин участвует в синтезе амикислот - цистеина (гомоцистеин + серин). Креатин подвергается фосфорилированию с участием АТФ, в результате образуется соединение с макроэргической связью - креатинфосфат. Это обратимая реакция, которая катализируется ферментом креатинфосфокиназой (КФК).
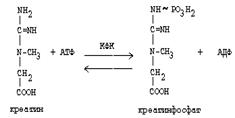
|
Эта реакция интенсивно идет в мышцах, особенно, в сердечной мышце, и в ткани мозга. Креатинфосфат активно синтезируется в покое и распадается при мышечной работе. Это наиболее быстрый способ регенерации АТФ. Креатин, образовавшийся из креатинфосфата распадается до креатинина, который является конечным продуктом и выводится с мочой. В сутки выводится 1-2 грамма креатинина. Это количество креатинина прямо пропорционально мышечной массе, поэтому у мужчин креатинина в моче больше, чем у женщин. Креатинин не реабсорбируется из первичной мочи, поэтому его количество во вторичной моче характеризует объем клубочковой фильтрации.
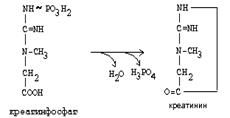
|
При поражении мышечных клеток и нарушении ткани мозга креатинфосфокиназа появляется в крови, это является диагностическим признаком. Известно, что КФК имеет три изофермента - кардиальный, церебральный и мышечный, появление их в крови позволяет определить поражение соответствующего органа. Появление в крови кардиального изофермента является ранним диагностическим признаком инфаркта миокарда.
ОБМЕН ЦИСТЕИНА.
Это заменимая аминокислота, она синтезируется из серина, гидроксильная группа которого замещается SН-группой, которую поставляет гомоцистеин. Цистеин в составе белков-ферментов своей -SН группой участвует в образовании каталитического центра (тиоловые протеазы), а также участвует в образовании дисульфидных связей, которые принимают участие в формировании третичной и четвертичной структуры белков. Также цистеин необходим для синтеза трипептида глютатиона, который состоит из цистеина и глютаминовой кислоты. Молекула глутатиона условно обозначается как (Г-SН). Глютатион способен легко окисляться и восстанавливаться:
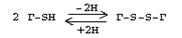
|
Окисляясь, глютатион предохраняет от окисления другие вещества, например, двухвалентное железо гемоглобина в эритроцитах:

|
Восстанавливается глютатион с помощью НАДФН2 с участием фермента глютатионредуктазы.

Цистеин подвергается и распаду, при этом он окисляется и декарбоксилируется, в результате образуется таурин, который участвует в образовании парных желчных кислот (таурохолевая и др.) в печени.
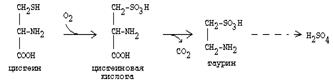
|
Серная кислота, которая образуется из таурина, участвует в обезвреживании токсических веществ в печени.
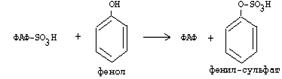
|
Так обезвреживаются продукты гниения белков в кишечнике - индол, скатол, фенол и крезол. В этих процессах серная кислота участвует в своей активной форме в виде 3’-фосфоаденозин-5’-фосфосульфата (ФАФC), которая образуется с участием АТФ.
ОБМЕН АРГИНИНА
Аргинин - частично незаменимая аминокислота. Она образуется в ходе синтеза мочевины в печени из карбомоилфосфата при участии аспарагиновой кислоты и орнитина. Аргинин участвует в синтезе креатина в почках, являясь донором гуанидиновой группы в образовании гуанидинацетата. В составе белков аргинин как полярная положительно заряженная аминокислота участвует в образовании ионных связей и в формировании гидратной оболочки белков.
ОБМЕН ДИКАРБОНОВЫХ АМИНОКИСЛОТ - ГЛУТАМИНОВОЙ И АСПАРАГИНОВОЙ
Эти аминокислоты вступают в многочисленные химические реакции, с которыми Вы уже знакомы и поэтому они играют главную роль в обмене аминокислот.
1. Они участвуют в реакциях синтеза заменимых аминокислот и следовательно в коррекции аминокислотного состава белков, а, значит, в коррекции аминокислотного состава клеток организма.
2. Участвуют в реакциях обезвреживания аммиака и других токсичных продуктов азотистого обмена.
3. Превращаясь в альфа-кетокислоты (альфакетоглутарат и ЩУК), они принимают участие во взаимосвязи обмена белков с обменом углеводов и жиров.
4. Дикарбоновые аминокислоты и их амиды (глутамин и аспарагин) участвуют в реакциях синтеза почти всех азотсодержащих соединений клеток (нуклеотидов, нуклеиновых кислот, аминосахаров и аминопроизводных липидов). В этих реакциях синтеза они являются донором азота в виде NН2-группы, или участвуют всей своей молекулой.
КОНКРЕТНАЯ РОЛЬ КАЖДОЙ АМИНОКИСЛОТЫ:
Глутаминовая кислота
а) подвергается прямому окислительному дезаминированиюс образованием альфа-кетоглутарата,
б) вступает в реакции трансаминирования, которые катализируют специфические трансаминазы,
в) является субстратом для синтеза глютамина, который является транспортной формой аммиака и участвует в синтезе мочевины в печени, также глютамин участвует в синтезе пуриновых оснований нуклеотидов и нуклеиновых кислот, аминосахаров и аминопроизводных липидов.
г) принимает участие в косвенном дезаминировании АК-т,
д) участвует в синтезе трипептида глютатиона,
е) является субстратом для образования гамма- аминомасляной кислоты.
Аспарагиновая кислота
а) участвует в реакциях трансаминирования,
б) в синтезе мочевины, как донор NН2-группы,
в) в синтезе пиримидиновых оснований (всей молекулой) и как донор NН2-группы - в синтезе пуриновых оснований,
г) в синтезе аспарагина.
Обе аминокислоты участвуют в формировании третичной и четвертичной структур белков, так как являются полярными заряженными аминокислотами и в образовании гидратной оболочки белков, а также эти аминокислоты принимают участие в формировании активных центров ферментов. Амидирование аспарагиновой и глутаминовой кислот приводит к образованию глутамина и аспарагина, необходимых для синтеза белков.
ОБМЕН ЦИКЛИЧЕСКИХ АМИНОКИСЛОТ ФЕНИЛАЛАНИНА И ТИРОЗИНА
Фенилаланин является незаменимой аминокислотой, а тирозин - заменимая аминокислота.
Фенилаланин вступает в незначительное количество превращений в тканях. Кроме включения этой аминокислоты в структуру молекул белка, единственным путем метаболизма фенилаланина у здорового человека является его окисление в тирозин с участием фермента микросомального окисления, специфической монооксигеназой - фенилаланингидроксилазой:
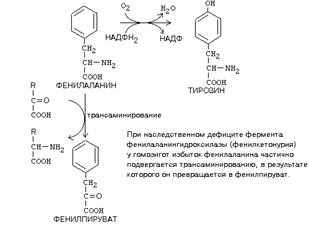
|
Тирозин вступает в многочисленные реакции в различных тканях. В результате этих превращений тирозин не только распадается до конечных продуктов, но и дает промежуточные метаболиты, из которых образуются ряд важных соединений, некоторые из которых являются биологически активными веществами.
Из тирозина образуются:
а) гормоны мозгового слоя надпочечников адреналин и норадреналин,
б) меланины - пигменты кожи, волос, радужной оболочки глаза,
в) йодсодержащие гормоны щитовидной железы - тироксин и трийодтиронин.
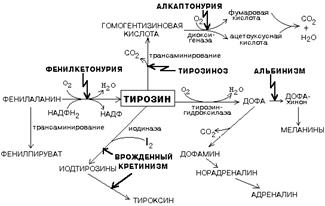
НАРУШЕНИЯ ОБМЕНА ФЕНИЛАЛАНИНА И ТИРОЗИНА
Нарушения обмена этих АК связано с нарушением биосинтеза некоторых ферментов, которые катализируют метаболические превращения этих АК. Результатом нарушения синтеза ферментов является возникновение наследственных генетических заболеваний:
1) фенилкетонурия - нарушен синтез фенилаланин-гидроксилазы, поэтому фенилаланин превращается в фенилпируват, который оказывает токсическое воздействие на развитие некоторых отделов головного мозга.
2) альбинизм - нарушен синтез ферментов, превращающих ДОФА в ДОФА-хром, поэтому нарушается синтез меланинов.
3) алкаптонурия - нарушен синтез диоксигеназы гомогентизиновой кислоты, она выделяется с мочой, моча приобретает черный цвет.
4) кретинизм - нарушен синтез йодиназы, что приводит к нарушению синтеза йодсодержащих гормонов щитовидной железы.
5) может быть нарушен синтез фермента тирозиназы, который катализирует превращение тирозина в ДОФА, следовательно будет нарушаться синтез гормонов мозгового слоя надпочечников и меланина.
Из всех этих заболеваний в настоящее время удается лечить фенилкетонурию, для этого из рациона ребенка исключают фенилаланин и увеличивают в пище количество тирозина. Если ребенка держать на этой диете до 6-7 лет, тогда не возникает умственная отсталость, т.к. к 6-7 годам успевают развиться отделы головного мозга, развитие которых задерживается при избытке в ткани мозга фенилпирувата.
Дата добавления: 2019-04-03; просмотров: 701;