И ИХ ПЕРЕНОС КРОВЬЮ
Переход О  из альвеолярного воздуха в кровь и СО из крови в альвеолы происходит только путем диффузии. Никакого механизма активного транспорта газов здесь не существует. Движущей силой диффузии являются разности (градиенты) парциальных давлений (напряжений) О и СО по обе стороны альвеолярно-капиллярной мембраны или аэрогематического барьера Напряжение газов в различных средах представлено в таблице 5.
из альвеолярного воздуха в кровь и СО из крови в альвеолы происходит только путем диффузии. Никакого механизма активного транспорта газов здесь не существует. Движущей силой диффузии являются разности (градиенты) парциальных давлений (напряжений) О и СО по обе стороны альвеолярно-капиллярной мембраны или аэрогематического барьера Напряжение газов в различных средах представлено в таблице 5.
Кислород и углекислый газ диффундируют только в растворенном состоянии, что обеспечивается наличием в воздухоносных путях водяных паров, слизи и сурфактантов. В ходе диффузии через аэроге-матический барьер молекулы растворенного газа преодолевают большое сопротивление, обусловленное слоем сурфактанта, альвеолярным эпителием, мембранами альвеол и капилляров, эндотелием сосудов, а также плазмой крови и мембраной эритроцитов.
Диффузионная способность легких для кислорода очень велика. Это обусловлено огромным числом (сотни миллионов) альвеол и
Таблица 5
Напряжение О и СО (мм рт. ст.) при спокойном дыхании воздухом
| Среда | Кислород | Углекислый газ |
| Альвеолярный воздух Венозная кровь Артериальная кровь Ткани | 10-20 | 50-60 |
большой их газообменной поверхностью (около 100 м 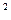 ), а также малой толщиной (около 1 мкм) альвеолярно-капиллярной мембраны. Диффузионная способность легких у человека примерно равна 25 мл О в 1 мин в расчете на 1 мм рт. ст. градиента парциальных давлений кислорода. Учитывая, что градиент рО между притекающей к легким венозной кровью и альвеолярным воздухом составляет около 60 мм рт. ст., этого оказывается достаточно, чтобы за время прохождения крови через легочный капилляр (около 0.8 с) напряжение кислорода в ней успело уравновеситься с альвеолярным рО .
), а также малой толщиной (около 1 мкм) альвеолярно-капиллярной мембраны. Диффузионная способность легких у человека примерно равна 25 мл О в 1 мин в расчете на 1 мм рт. ст. градиента парциальных давлений кислорода. Учитывая, что градиент рО между притекающей к легким венозной кровью и альвеолярным воздухом составляет около 60 мм рт. ст., этого оказывается достаточно, чтобы за время прохождения крови через легочный капилляр (около 0.8 с) напряжение кислорода в ней успело уравновеситься с альвеолярным рО .
Диффузия СО из венозной крови в альвеолы даже при сравнительно небольшом градиенте рСО (около 6 мм рт. ст.) происходит достаточно легко, так как растворимость СО в жидких средах в 20-25 раз больше, чем у кислорода. Поэтому после прохождения крови через легочные капилляры рСО в ней оказывается равным альвеолярному и составляет около
40 мм рт. ст.
Дыхательная функция крови прежде всего обеспечивается доставкой к тканям необходимого им количества О Кислород в крови находится в двух агрегатных состояниях: растворенный в плазме (0.3 об.%) и связанный с гемоглобином (около 20 об.%) — оксигемоглобин.
Отдавший кислород гемоглобин считают восстановленным или дезоксигемоглобином. Поскольку молекула гемоглобина содержит 4 частицы гема (железосодержащего вещества), она может связать четыре молекулы О . Количество О , связанного гемоглобином в 100 мл крови, носит название кислородной емкости крови и составляет около 20 мл О . Кислородная емкость всей крови человека, содержащей примерно 750 г гемоглобина, приблизительно равна 1 л.
Каждому значению рО в крови соответствует определенное процентное насыщение гемоглобина кислородом. Кривую зависимости процентного насыщения гемоглобина кислородом от величины парциального напряжения называют кривой диссоциации оксигемоглобина (рис. 21). Анализ хода этой кривой сверху вниз показывает, что с уменьшением рО в крови происходит диссоциация оксигемоглобина, т. е. процентное содержание
оксигемоглобина уменьшается, а восстановленного растет.
В различных условиях деятельности может возникать острое снижение насыщенности крови кислородом —гипоксемия. Причины гипоксемии весьма разнообразны. Она может развиваться вследствие снижения рО в альвеолярном воздухе (произвольная задержка дыхания, вдыхание воздуха с пониженным рО ), при физических нагрузках, а также при неравномерной вентиляции различных отделов легких.
Образующийся в тканях СО диффундирует в тканевые капилляры, откуда переносится венозной кровью в легкие, где переходит в
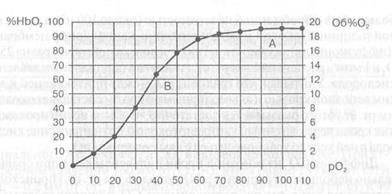
Рис. 21. Кривая диссонации оксигемоглобина в крови человека в покое
А — содержание НbO в артериальной крови, В — то же в венозной крови
альвеолы и удаляется с выдыхаемым воздухом. Углекислый газ в крови (как и О ) находится в двух состояниях: растворенный в плазме (около 5% всего количества) и химически связанный с другими веществами (95%). СО в виде химических соединений имеет три формы: угольная кислота (Н СО  ), соли угольной кислоты (NaHCO ) ив связи с гемоглобином (НвНСО ).
), соли угольной кислоты (NaHCO ) ив связи с гемоглобином (НвНСО ).
В крови тканевых капилляров одновременно с поступлением СО внутрь эритроцитов и образованием в них угольной кислоты происходит отдача О оксигемоглобином. Восстановленный Н в легко связывает водородные ионы, образующиеся при диссоциации угольной кислоты. Таким образом, восстановленный Н в венозной крови способствует связыванию СО , аоксигемоглобин, образующийся в легочных капиллярах, облегчает его отдачу.
В состоянии покоя с дыханием из организма человека удаляется 230-250 мл СО в 1 минуту. При удалении из крови СО из нее уходит примерно эквивалентное число ионов водорода.
Таким порядком дыхание участвует в регуляции кислотно-щелочного состояния во внутренней среде организма.
Обмен газов между кровью и тканями осуществляется также путем диффузии. Между кровью в капиллярах и межтканевой жидкостью существует градиент напряжения О , который составляет 30-80 мм рт. ст., а напряжение СО в интерстициальной жидкости на 20-40 мм рт. ст. выше, чем в крови. Кроме того, на обмен О и СО в тканях влияют площадь обменной поверхности, количество эритроцитов в крови, скорость кровотока, коэффициенты диффузии газов в тех средах, через которые осуществляется их перенос.
Артериальная кровь отдает тканям не весь О . Разность между об.% О в притекающей к тканям артериальной крови (около 20 об.%) и оттекающей от них венозной кровью (примерно 13 об.%)
называется артерио-венозной разностью по кислороду (7 об.%). Эта величина служит важной характеристикой дыхательной функции крови, показывая, какое количество О доставляют тканям каждые 100 мл крови. Для того, чтобы установить, какая часть приносимого кровью О, переходит в ткан и, вычисляют коэффициент утилизации (использования) кислорода. Его определяют путем деления величины артерио-венозной разности на содержание О в артериальной крови и умножения на 100. В покое для всего организма коэффициент утилизации О равен примерно 30-40%. Однако в миокарде, сером веществе мозга, печени и корковом слое почек он составляет 40-60%. При тяжелых физических нагрузках коэффициент утилизации кислорода работающими скелетными мышцами и миокардом достигает 80-90%.
В снабжении мышц О при тяжелой работе имеет определенное значение внутримышечный пигмент м и о г л о б и н, который связывает дополнительно 1.0-1.5.Л О . Связь О с миоглобином более прочная, чем с гемоглобином. Оксимиоглобин отдает О только при выраженной гипоксемии.
РЕГУЛЯЦИЯ ДЫХАНИЯ
Регуляция внешнего дыхания представляет собой физиологический процесс управления легочной вентиляцией для обеспечения оптимального газового состава внутренней среды организма в постоянно меняющихся условиях его жизнедеятельности. Основную роль в регуляции дыхания играют рефлекторные реакции, возникающие в результате возбуждения специфических рецепторов, заложенных в легочной ткани, сосудистых рефлексогенных зонах и скелетных мышцах. Центральный аппарат регуляции дыхания представляют нервные образования спинного, продолговатого мозга и выше лежащих сегментов ЦНС.
Гуморальная регуляция дыхания, созданная Д. Холденом и Д. Пристли около 50 лет тому назад, в последние годы не находит экспериментального подтверждения, большинством специалистов считается ошибочной и упоминается сейчас только в историческом плане. Это обусловлено открытием специфических рецепторов (механо- и хеморецепторов), а также других рефлекторных влияний на дыхательный центр. Поэтому все изменения внешнего дыхания в настоящее время объясняются только рефлекторными механизмами.
Дыхательный ритм и управление деятельностью дыхательных мышц генерируется работой дыхательного центра, представляющего собой совокупность взаимосвязанных нейронов ретикулярной формации продолговатого мозга и вышележащих отделов ЦНС, обеспечивающих тонкое приспособление дыхания
к различным условиям внешней среды. Современные представления о работе дыхательного центра сводятся к тому, что часть дыхательных нейронов, объединенных в так называемую латеральную зону является эфферентной частью дыхательного центра и обеспечивает преимущественно фазу вдоха (инспираторные нейроны). Другая группа нейронов, составляющая медиальную зону, является афферентной частью дыхательного центра и обеспечивает фазу выдоха (экспираторные нейроны). Предназначение этой зоны заключается в контроле за периодичностью дыхательной ритмики, организуемой латеральной зоной.
В регуляции дыхания на основе механизма обратных связей принимают участие несколько групп механорецепторов легких.
Рецепторы растяжения легких находятся в гладких мышцах трахеи и бронхов. Адекватным раздражителем этих рецепторов является растяжение стенок воздухоносных путей.
Ирритантные рецепторы расположены в эпителиальном слое верхних дыхательных путей и раздражаются при изменении объема легких, а также при пневмотораксе, коллапсе и действии на слизистую трахеи и бронхов механических или химических раздражителей. При раздражении этих рецепторов у человека возникают кашлевой рефлекс, першение и жжение, учащение дыхания и броноспазм.
Джи - рецепторы расположены в стенках альвеол в местах их контакта с капиллярами, поэтому их еще называют юкстакапиллярные рецепторы легких. Эти рецепторы формируют частое поверхностное дыхание при патологии легких (воспаление, отек, повреждения легочной ткани), а также раздражаются при действии некоторых биологически активных веществ (никотин, гистамин и др.).
Проприорецепторы дыхательных мышц (межреберные мышцы, мышцы живота) обеспечивают усиление вентиляции легких при повышении сопротивления дыханию.
Поддержание постоянства газового состава внутренней среды организма регулируется с помощью центральных и периферических хеморецепторов.
Центральные хеморецепторы расположены в структурах продолговатого мозга, и они чувствительны к изменению рН межклеточной жидкости мозга. Эти рецепторы стимулируются ионами водорода, концентрация которых зависит от рСО в крови. При снижении рН интерстициальной жидкости мозга (концентрация водородных ионов растет) дыхание становится более глубоким и частым. Напротив, при увеличении рН угнетается активность дыхательного центра и снижается вентиляция легких.
Периферические (артериальные) хеморецепторы расположены в дуге аорты и месте деления общей сонной артерии (каротидный синус). Эти рецепторы вызывают рефлекторное
увеличение легочной вентиляции в ответ на снижение рО в крови
(гипоксемия).
Афферентные влияния с работающих мышц осуществляются благодаря раздражению проприорецепторов, что приводит к усилению дыхания рефлекторным путем. Повышение активности дыхательного центра в этом случае является результатом распространения возбуждения по различным отделам ЦНС.
Существенное воздействие на регуляцию дыхания оказывают и условнорефлекторные влияния. В частности, эмоциональные нагрузки, предстартовые состояния, гипнотические внушения, влияния индифферентных раздражителей, сочетавшихся ранее с избытком СО , самообучение управлению дыханием подтверждают сказанное. Легочная вентиляция зависит также от особенностей гемодинамики (уровень АД, величина МОК), температуры внешней среды и других факторов.
ПИЩЕВАРЕНИЕ
Пищеварением называется процесс физической и химической переработки пищи, в результате которого становится возможным всасывание питательных веществ из пищеварительного тракта, поступление их в кровь и лимфу и усвоение организмом.
Дата добавления: 2017-09-19; просмотров: 327;