Всмоктування вуглеводів.
Всмоктування — це складний біохімічний процес переходу молекул моносахаридів та їхніх ефірів через епітелій слизової оболонки тонкої кишки у кров і лімфу,
Деяка кількість моносахаридів (до 10 %) всмоктується слизовою оболонкою шлунка. Тонка кишка має величезну всмоктувальну поверхню (у людини вона досягає 500 м2). Збільшенню такої поверхні сприяє наявність у слизовій оболонці ворсинок (2—2,5 тис. на 1 см2) та мікроворсинок (2—3 тис. на 1 клітину). Кормові маси тут перебувають в середньому 8—9 год. За такий проміжок часу практично всі моносахариди, що містяться в кормі, всмоктуються через слизову оболонку в кров.
Пентози всмоктуються повільніше, ніж гексози. Гексози всмоктуються переважно у вигляді гексозофосфатів, що перешкоджає вирівнюванню осмотичної концентрації в епітелії в міру всмоктування і забезпечує безперервне надходження в кров'яне русло гексоз. На швидкість всмоктування впливає структура молекули гексози. Після переходу через кишкову стінку гексозофосфати гідроліуються: моносахариди надходять у кровотік, Н3Р04 використовується для фосфорилування нових порцій вуглеводів, які всмоктуються. Під час всмоктування частина моносахаридів (фруктоза, галактоза, маноза) таутомеризується в глюкозу.
Механізм всмоктування пояснює теорія переносників, згідно з якою транспортування моносахаридів у клітину епітелію ворсинки здійснюється за допомогою спеціальних білків з молекулярною масою 10—30 тис. Вони знаходяться на мембранах мікроворсинок і відразу ж після ферментативного розщеплення дисахаридів сприймають на певні «площадки» своїх молекул моносахариди, які потім переносять вглиб клітини до базального її кінця.
Переносник сполучається з моносахаридами та їхніми дериватами. Мітохондрії є в цих процесах джерелом хімічної енергії. Комплекс переносник — моносахарид може пересуватися вглиб клітини по ендоплазматичній сітці та інших органоїдах. На базальній поверхні клітини комплекс розпадається. Переносник повертається до поверхні клітини і сполучається з новими порціями моносахаридів. Моносахарид потрапляє в міжклітинну рідину, потім у судинну систему — капіляри, підепітеліальну і підслизову венозну сітку, вени брижі і ворітну вену.
Процес активується іонами Na+, які створюють натрієвий насос, що забезпечує активне транспортування моносахаридів через мембрани ентероцита
У товстій кишці (сліпа, ободова, пряма) частина полісахаридів корму, яка не зазнала гідролізу, розкладається під впливом мікробів. Продукти, що при цьому утворилися, всмоктуються слизовою оболонкою в кров'яне русло. При недоброякісному годуванні і деяких захворюваннях травного каналу утворюються продукти гниття, які чинять на організм токсичну дію. У товстій кишці травоїдних тварин деякі мікроорганізми синтезують вітаміни (К, В1, В2, В6, Вз, В5, С).
Проміжний обмін. Складається з двох процесів — біосинтезу і розпаду. Він відбувається в органах, тканинах, клітинах та інтрацелюлярних структурах. При цьому моносахариди крові використовуються для різних потреб. Так, у людини 3—5 % глюкози крові використовується для синтезу глікогену, 30—35 — для синтезу ліпідів, 60—70 % є джерелом хімічної енергії. Проміжний обмін часто називають внутрішньотканинним або внутрішньоклітинним обміном.
При розщепленні молекули вуглеводу до СО2 і Н2О в тканинах і клітинах утворюється велика кількість інших проміжних продуктів обміну.
Глюкоза знаходиться у вільному і зв'язаному стані. Кількість зв'язаної глюкози досягає 40—50 % загального її вмісту в крові. Артеріальна кров, яка надходить у головний мозок по сонних артеріях, на 5,44—5,83 ммоль/л багатша на глюкозу, ніж венозна, що відтікає від мозку по яремних венах.
У тканинах і клітинах основними енергетичними перетвореннями вуглеводів є: анаеробне розщеплення, цикл трикарбонових кислот Кребса і пентозофосфатний шлях (ПФШ), або пентозний шлях. Усі три процеси взаємозв'язані, оскільки в кожному з них є спільні для всього проміжного обміну вуглеводів продукти хімічних реакцій і беруть участь одні й ті самі ферментні системи.
Анаеробне розщеплення вуглеводів. Цей процес відбувається в органах, тканинах і клітинах тваринного організму без участі кисню. Його основні реакції подібні до хімізму спиртового бродіння, названого Л. Пастером «життям без кисню». Відмінності полягають ось у чому: під час спиртового бродіння вуглеводів кінцевими продуктами є етанол і СО2, а при анаеробному розщепленні — молочна або піровиноградна кислота.
Анаеробне розщеплення вуглеводів може починатися фосфоро-ітичним розщепленням глікогену (глікогеноліз) або фосфорилуванням глюкози (гліколіз). У скелетних м'язах обидва процеси виражені однаковою мірою, а в головному мозку і міокарді переважає гліколіз.
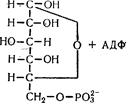
Фосфоролітичне розщеплення глікогену до глюкозо-1-фосфату відбувається під впливом ферменту фосфорилази:
 H-C-O-PO32-
H-C-O-PO32-
 H-C-OH
H-C-OH
 HO-C-H
HO-C-H
 (C6H10O5)n+nH3PO4 ® H-C-OH
(C6H10O5)n+nH3PO4 ® H-C-OH
 H-C
H-C
CH2OH
Під впливом ферменту фосфоглюкомутази глюкозо-1-фосфат перетворюється на глюкозо-6-фосфат.
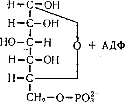
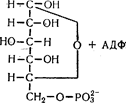
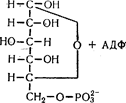
Якщо процес починається з глюкози, то відбувається її фосфорилування під впливом ферменту фосфоглюкокінази з утворенням глюкозо-6-фосфату:

|
Після цього подальші шляхи глікогенолізу і гліколізу зливаються і можуть бути розглянуті разом.
1. Глюкозо-6-фосфат під дією ферменту глюкозофосфатізомерази перетворюється на фруктозо-6-фосфат. Між обома ефірами встановлюється рівновага при вмісті 70 % глюкозо-6-фосфату і 30 % фруктозо-6-фосфату:
   2. Під впливом фосфофруктокінази, при наявності АТФ та іонів Мg2+ утворюється фруктозо-1,6-дифосфат
2. Під впливом фосфофруктокінази, при наявності АТФ та іонів Мg2+ утворюється фруктозо-1,6-дифосфат
|
|
3. Наявність двох залишків фосфату на протилежних кінцях молекули гексози призводить до різкого ослаблення зв'язків між третім і четвертим атомами вуглецю. В результаті цього вуглевод легко розщеплюється на дві фосфотріози під впливом ферменту альдолази:
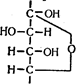
|
При цьому утворюється 3-фосфогліцериновий альдегід (3 %) і діоксіацетонфосфат (97 %), який таутомеризується у 3-фосфогліцериновий альдегід:
4. 3-фосфогліцериновий альдегід під впливом ферменту гліце-ральдегідфосфатдегідрогенази і НАД вступає в реакцію оксиредукції, що приводить до утворення макроергічного зв'язку, а потім 1,3-дифосфогліцеринової кислоти:
5. 1,3-Дифосфогліцеринова кислота під впливом ферменту фосфо-гліцераткінази віддає свій макроерп'чний зв'язок АДФ:
6. 3-Фосфогліцеринова кислота під впливом ферменту фосфоглі-церомутази перетворюється на 2-фосфогліцеринову кислоту:
7. 2-Фосфогліцеринова кислота під впливом ферменту фосфопіру-ватгідратази при наявності іонів Мg2+ втрачає молекулу води, перетворюючись на енольну форму фосфопіровиноградної кислоти, що містить макроергічний зв'язок:
8. Фосфопіровиноградна кислота під впливом ферменту піруваткіна-зи при наявності іонів М^24" і К+ віддає свій макроергічний зв'язок АДФ, перетворюючись на енольну форму піровиноградної кислоти:
Енольна форма піровиноградної кислоти таутомеризується в кетоформу:
9. При нестачі кисню піровиноградна кислота під дією ферменту лактатдегідрогенази і при наявності НАД • Н + Н+ перетворюється на молочну кислоту, яка звичайно є кінцевим продуктом анаеробного розпаду вуглеводів у тваринних тканинах;
Отже, під час анаеробного окислення з однієї молекули глюкози утворюється чотири молекули АТФ. Якщо врахувати, що на фосфорилування глюкози і утворення фруктозо-1-дифосфату витрачається дві молекули АТФ, чистий вихід енергії дорівнюватиме двом молекулам АТФ. При глікогенолізі з однієї молекули глюкози утворюється три молекули АТФ. При гліколізі або глікогенолізі в АТФ акумулюється близько 35—40 % всієї вивільнюваної хімічної енергії. Решта 60—65 % енергії розсіюється у вигляді теплоти.
Отже, коефіцієнт корисної дії анаеробного гліколізу клітин чи тканин дорівнює 0,35—0,40.
Цикл трикарбонових кислот Кребса (ЦТК).Основна маса хімічної енергії вуглеводу вивільнюється в аеробних умовах за участю кисню. Цикл трикарбонових кислот Кребса називають ще циклом лимонної кислоти, або клітинним диханням.
Між анаеробним і аеробним розщепленням вуглеводів існує тісний зв'язок. Перш за все він виявляється у наявності піровиноградної кислоти, якою завершується анаеробне розщеплення вуглеводів і починається клітинне дихання. Обидві фази каталізуються одними й тими самими ферментами. Хімічна енергія, що вивільнюється під час фосфорилування, резервується у вигляді макроергів АТФ. У хімічних реакціях беруть участь одні й ті самі коферменти (НАД, НАДФ) і катіони. Відмінності полягають ось у чому: якщо анаеробне розщеплення вуглеводів переважно локалізоване в гіалоплазмі, то реакції клітинного дихання відбуваються в основному в мітохондріях.
За деяких умов виявляється антагонізм між обома фазами. Так, при наявності кисню швидкість реакції гліколізу різко зменшується. Продукти гліколізу можуть гальмувати аеробний обмін вуглеводів
ЦТК є послідовним ланцюгом хімічних реакцій, в результаті яких продукти розщеплення вуглеводів окислюються до СО2 і Н2О, а хімічна енергія акумулюється в макроергічних сполуках. У ході клітинного дихання утворюється «носій» — щавлево-оцтова кислота (ЩОК). У подальшому відбувається конденсація з «носієм» активованого залишку оцтової кислоти. Виникає трикарбонова кислота — лимонна. У ході хімічних реакцій відбувається «оберт» залишку оцтової кислоти в циклі. З кожної молекули піровиноградної кислоти утворюється 18 молекул АТФ. У кінці циклу вивільнюється «носій», який вступає в реакцію з новими молекулами активованого залишку оцтової кислоти,
1. Якщо кінцевим продуктом анаеробного розщеплення вуглеводів є молочна кислота, то під впливом лактатдегідрогенази вона окислюється до піровиноградної кислоти:
2. Частина молекул піровиноградної кислоти йде на синтез «носія» ЩОК під впливом ферменту піруваткарбоксилази і при наявності іонів Мg2+
3. Частина молекул піровиноградної кислоти є джерелом утворення «активного ацетату» — ацетилкоензиму А (ацетил-КоА). Реакція відбувається під впливом піруватдегідрогенази:
Ацетил-КоА містить макроергічний зв'язок, у якому акумулюється близько 5—7 % енергії. Основна маса хімічної енергії утворюється в результаті окислення «активного ацетату».
4. Під впливом цитратсинтетази починає функціонувати власне цикл трикарбонових кислот, що приводить до утворення лимонної кислоти:
5. Лимонна кислота під впливом ферменту аконітат-гідратази дегідруеться і перетворюється на цис-аконітову кислоту, яка після приєднання молекули води переходить в ізолимонну:
Між трьома трикарбоновими кислотами встановлюється динамічна рівновага.
6. Ізолимонна кислота (окислюється до щавлево-янтарної, яка декарбоксилується і перетворюється на a-кетоглутарову кислоту. Реакція каталізується ферментом ізоцитратдегідрогеназою:
7. a-Кетоглутарова кислота під впливом ферменту 2-оксо-(a-кето)-глутаратдегідрогенази декарбоксилується, в результаті чого утворюється сукциніл-КоА, що містить макроергічний зв'язок:
8. На наступній стадії сукциніл-КоА під впливом ферменту сукци-ніл-КоА-синтетази передає макроергічний зв'язок ГДФ:
ГТФ під впливом ферменту ГТФ-аденілаткінази віддає макроергічний зв'язок АМФ:
9. Янтарна кислота під дією ферменту сукцинатдегідрогенази (СДГ) окислюється до фумарової. Коферментом СДГ є ФАД:
10. Фумарова кислота під впливом ферменту фумаратгідратази перетворюється на яблучну:
11. Яблучна кислота під впливом ферменту малатдегідрогенази (МДГ) окислюється, утворюючи ШОК:
При наявності в реагуючій системі ацетил-КоА ЩОК знову включається в цикл трикарбонових кислот.
Отже, з однієї молекули глюкози утворюється до 38 молекул АТФ (дві — за рахунок анаеробного гліколізу, шість — в результаті окислення двох молекул НАД • Н + Н+, що виникли під час гліколітичної оксиредукції, і ЗО — за рахунок ЦТК). Коефіцієнт корисної дії ЦТК дорівнює 0,5. Решта енергії розсіюється у вигляді теплоти. В ЦТК окислюється 16—33 % молочної кислоти, решта її маси йде на ресинтез глікогену.
Пентозний шлях. Пентозний шлях — це ланцюг послідовних хімічних перетворень вуглеводів, в результаті якого в тканинах і клітинах вивільнюється хімічна енергія і утворюються пентози, необхідні для синтезу нуклеїнових кислот, нуклеотидів і коферментів. Його часто називають апотомічним шляхом, оскільки під час окислення глюкози відщеплюється один атом вуглецю. Іноді його називають прямим, або гексозомонофосфатним, шляхом окислення вуглеводів, оскільки тут глюкозо-6-фосфат піддається прямому окисленню (з відщепленням СОз) без утворення фруктозо-1,6-дифосфату і двох фосфотріоз. Пентозний шлях складається з таких стадій:
1. Глюкозо-6-фосфат, що утворюється в результаті фосфоролізу глікогену або фосфорилування глюкози, під впливом глюкозо-6-фос-фатдегідрогенази окислюється і перетворюється на 6-монофосфат-глюкон:
2. 6-Монофосфоглюкон під впливом ферменту глюконолактонази приєднує молекулу води, перетворюючись на 6-фосфоглюконову кислоту:

|
3. 6-Фосфоглюконова кислота за участю фосфоглюконатдегід-рогенази піддається окислювальному декарбоксилуванню, що приводить до утворення кетопентози — D-рибулозо-5-фосфату і другої молекули НАДФ • Н + Н+:
4. D – Рибулозо –5-фосфат під впливом рибулозофосфат –3 епімерази перетворюється на свій епімер – D- ксилулозо-5 фосфат
У деяких випадках D-рибулозо-5-фосфат може обернено перетворюватись на свій альдоізомер — D-рибозо-5-фосфат:
D-Рибозо-5-фосфат використовується клітинами для синтезу РНК і нуклеотидів (наприклад, АМФ, АДФ, АТФ). Часто пентозний шлях на даній стадії завершується. Його можна подати сумарним рівнянням:
Глюкозо-6-фосфат + 2НАДФ+Û D-Рибозо-5-фосфат + СО2 + +2НАДФ • Н+2Н+.
Пентози, не використані для синтезу нуклеїнових кислот і нуклеотидів, витрачаються на біосинтез інших сполук, регенерацію глюкози, з якої починався пентозний шлях.
5. Частина D-ксилулозо-5-фосфату та D-рибозо-5-фосфату під впливом транскетолази вступає в неокислений етап, що приводить до утворення D-седогептулозо-7-фосфату і 3-фосфогліцеринового альдегіду
6. Фосфогліцериновий альдегід може включатися в четверту стадію анаеробного розщеплення вуглеводів або під впливом ферменту трансальдолази взаємодіяти з D-седогептулозо-7-фосфатом, утворюючи фруктозо-6-фосфат і еритрозо-4-фосфат:
Фруктозо-6-фосфат може ізомеризуватися в глюкозо-6-фосфат і вступати в першу стадію пентозного шляху або включатися в другу стадію анаеробного розщеплення вуглеводів.
7. Еритрозо-4-фосфат під впливом ферменту транскетолази взаємодіє з ксилулозо-5-фосфатом, що приводить до утворення фруктозо-6-фосфату і 3-фосфогліцеринового альдегіду:
Реакції пентозного шляху можна підсумувати рівнянням:
6 Глюкозо-6-фосфат 4- 12НАДФ+ ® бСО2 + 12НАДФ • Н + Н+ +
+ 5 Глюкозо-6-фосфат + НзРО4.
Пентозний шлях відіграє важливу роль у синтезі жирів. Так, у жировій тканині він займає 50 % відносно гліколізу, в печінці — 2,5—3 і в м'язовій тканині—0,3 %. Припускають, що останні стадії пентозного шляху забезпечують жирові клітини гліцерином, який утворюється з 3-фосфогліцеринового альдегіду. Крім описаних вище в тканинах тваринного організму виявлені інші шляхи, зокрема перетворення моносахаридів шляхом їх сполучення з піримідиновими основами
Кінцевий обмін. Основними кінцевими продуктами обміну вуглеводів є вода і вуглекислий газ. Вода виділяється з сечею, потом, видихуваним повітрям, каловими масами, вуглекислий газ — через легені.
У сечі міститься деяка кількість глюкуронової кислоти, яка, знешкоджуючи отруйні речовини, утворює парні сполуки.
У калі міститься частина неперетравлених вуглеводів корму.
Регуляція вуглеводного обміну. В регуляції вуглеводного обміну беруть участь нервова система, залози внутрішньої секреції, печінка і вітаміни. Центри розміщені в корі великих півкуль, проміжному і довгастому мозку, гангліях вегетативної нервової системи. Фістульні досліди на залозах травного каналу свідчать про чільну роль ЦНС у регуляції вуглеводного обміну, оскільки її функціональним станом визначаються характер і інтенсивність секреції травних соків, ступінь глікогенезу і гліконеогенезу, швидкість реакцій глікогенолізу і гліколізу.
Існує прямий зв'язок між вмістом глюкози в крові і функціональним станом ЦНС. Так, зменшення концентрації глюкози в крові викликає збудження нервових центрів у гіпоталамусі і довгастому мозку. Нервові імпульси по аксонах через прикордонний симпатичний стовбур надходять у сонячне сплетіння, потім у печінку, де активізується фосфорилаза, яка розщеплює глікоген до глюкозо-1-фосфату. Рівень глюкози в крові відновлюється.
У регуляції вуглеводного обміну беруть участь гормони гіпофіза, коркової і мозкової речовин надниркових, підшлункової і щитовидної залоз. Так, гормон підшлункової залози інсулін, потрапляючи з током крові у печінку, активізує гексокіназу. Це приводить до утворення глюкозо-6-фосфату і глікогену, гальмує активність глюкозо-6-фосфатази. Глюкагон стимулює розпад глікогену шляхом активізації фосфорилази. Фосфюрилаза активізується гормонами надниркових залоз — адреналіном і норадреналіном. Під їхнім впливом відбувається розпад глікогену до глюкози.
Аналогічну дію має соматотропін, глюкокортикоїди коркового шару надниркових залоз і тироксин. Так, соматотропін гальмує фосфорилування глюкози і активізує інсуліназу. Глюкокортикоїди активізують глюкозо-6-фосфатазу, піруваткарбоксилазу, фосфопіру-ваткіназу, ферменти гліконеогенезу.
Посередником між гормонами і ферментами є цАМФ, яка «вмонтована» у мембрани клітин (формулу цАМФ див. на с. 95). Її діяльність активізується адреналіном, глюкагоном і АКТГ. Збільшується вміст цАМФ — зростає інтенсивність фосфоролізу глікогену. Між дією інсуліну, адреналіну, глюкагону та інших гормонів встановлюється динамічна рівновага, яка регулюється ЦНС.
Гепатоцити при високих концентраціях глюкози в крові можуть поглинаги надлишок вуглеводів, а при низьких — віддавати їх.
Майже всі вітаміни групи В беруть участь у регуляції вуглеводного обміну, особливо вітаміни Ві (складова частина ТПФ), РР (входить до складу НАДФ+), біотин (входить до складу піруваткарбоксилази), пантотенова кислота (складова частина коензиму А) тощо.
Патологія вуглеводного обміну. Найчастіше виявляється у вигляді гіперглікемії і глюкозурії, ацетонемії і ацетонурії, порушень азотистого, водного та мінерального обмінів тощо.
Гіперглікемія — вміст глюкози в крові вищий за норму. Вона призводить до глюкозурії — появи глюкози в сечі. Причиною їх може бути цукровий діабет — хвороба, що характеризується зниженим вмістом в організмі інсуліну, необхідного для перетворення глюкози на глікоген. У ряді випадків спостерігається гіпоглікемія — зменшення вмісту глюкози в крові нижче за норму. Причини цього можуть бути різні: підвищення вмісту інсуліну, зменшення інтенсивності синтезу антагоністів інсуліну, голодування, захворювання травного каналу тощо.
З порушеннями обміну вуглеводів пов'язані порушення обміну інших речовин, насамперед ліпідів (ще в минулому столітті з'явився крилатий вираз: жири згоряють у полум'ї вуглеводів). Вони виявляються у кетозах, ацетонемії, ацетонурії і коматозному стані.
Порушення процесів гліконеогенезу викликає надмірне руйнування білків і збільшення вмісту в крові та сечі продуктів азотистого обміну. Необхідність видалення з організму отруйних продуктів спричинює порушення водного і мінерального обмінів.
У ряді випадків (при цукровому діабеті) виникає поліурія — намірне виділення сечі і збіднення організму водою. З сечею «вимивається» багато мінеральних речовин.
Трапляються й інші порушення вуглеводного обміну: ідіопатична пентозурія (з сечею виділяється велика кількість пентоз), генетичного походження галактоземія і галактозурія, непереносність організмом лактози і сахарози, глікогенози тощо.
| <== предыдущая лекция | | | следующая лекция ==> |
| Обмін речовин і енергії | | | Структура дыхательной системы. |
Дата добавления: 2016-09-20; просмотров: 1260;