СТРАТИГРАФИЧЕСКОЕ РАСЧЛЕНЕНИЕ И СТРАТОТИПЫ. После установления меловой системы дальнейшая разработка ее стратиграфии была выполнена А
После установления меловой системы дальнейшая разработка
ее стратиграфии была выполнена А. д' Орбиньи, который широко
использовал палеонтологический метод. Взамен существующих
местных подразделений он ввел ярусы, каждый из которых характеризуется определенным фаунистическим комплексом.
Термин «неоком» был предложен в 1835 г. Дж. Турманном
для морских отложений нижней части меловой системы, развитых
в южных горах Швейцарии. Название происходит от древнеримского имени г. Невшателя. В дальнейшем неоком был утвержден
в качестве надъяруса и в его составе стали выделять 4 яруса.
Сенон впервые был обоснован в ранге яруса А. д' Орбиньи.
Название происходит от древнего имени г. Санса на р. Ионна. В
дальнейшем в составе сенона были выделены 4 яруса, а сам он
перешел в ранг надъяруса.
Стратотип берриасского яруса расположен на юго-востоке
Франции у д. Берриас. В 1871 г. известняки берриаса Г. Коканом
были выделены в самостоятельный ярус, и он поместил его в основание меловой системы. Позднее берриас был включен в качестве подъяруса в титонский ярус. С конца прошлого века развернулась дискуссия о статусе берриаса. Стратотипический разрез
переизучался в 60—70-е годы нашего столетия, и ряд исследователей предложили рассматривать берриас в качестве самостоятельного яруса.
Второй международный коллоквиум по границе юры и мела,
проведенный в Лионе и Невшателе в 1973 г., принял зональное
деление берриаса, утвердив его в качестве нижнего яруса меловой системы. Границу между титоном п берркасом предложено
проводить, как и раньше, в основании подзоны Pseudosubplamtcs
grandis.
Валанжинский ярус был выделен из неокома, развитого в
г. Невшатель (Швейцария), около замка Валанжин. К нему была
отнесена пачка переслаивания серых, голубых и желтых мергелей
и плотных органогенных известняков, в основании оолитовых, а в
верхах с железистыми оолитами. В толще, имеющей мощность
53—55 м, встречаются остатки морских ежей, брахиопод, губок,
мшанок, кораллов, фораминифер, двустворчатых и брюхоногих
моллюсков. В залегающем в кровле слое известково-мергелистых
желваков были найдены аммониты.
'Более поздние исследования показали, что значительная часть
выделенных слоев относится к берриасу, а сам стратотип валанжика выбран неудачно, так как разрез изобилует конденсированными слоями и перерывами. Крайне редко встречаются аммониты — обитатели пелагиали, но многочисленны представители бентоса. В решении Лионского коллоквиума (1963) предла1алось
найти и описать гипостратотип. В 1979 г. французские палеонтологи описали гипостратотип валанжина в Воконтской впадине
(юго-восточная Франция) около д. Англе, представленный относительно равномерным чередованием мергелей, глин и известняков, отлагавшихся в пелагической зоне моря, без видимых стратиграфических перерывов, и заключающих многочисленные остатки
аммонитов. Мощность отложений валанжина в гипостратотипе
составляет 244 м. На основании распределения аммонитов выделены два подъяруса, каждый из которых состоит из трех зон.
В 1873 г. Э. Реневье выделил в готеривский ярус отложения,
развитые у д. Отрив (Швейцария), расположенной около г. Невшателя. Они представлены мергелями и оолитовыми известняками
с остатками аммонитов, брахиопод, устриц и морских ежей. Позднее эти отложения на основании распределения аммонитов были
разделены на два подъяруса с двумя зонами в каждом.
Типичный разрез барремского яруса находится у д. Баррем
(юго-восточная Франция, бассейн р. Дюранс), где в известняках
обнаружены развернутые аммониты Ancyloceras, Scaphites
и др.
Аптский ярус впервые выделил А. д' Орбиньи среди отложении,
развитых у д. Апт в юго-восточной Франции. Он отнес к апту гли-
ны с Plicatula, а позднее и известняки с большим числом аммонитов. До сих пор нижняя и верхняя границы аптского яруса
дискуссионны. Ныне аптский ярус выделяется в количестве трех
подъярусов: нижний (бедуль), средний (гаргаз) и верхний (клансей). Название подъярусам даны по местностям, где развиты
стратотипические разрезы. Все подъярусы и зоны аптского яруса
охарактеризованы большим количеством аммонитов.
Альбский ярус выделил А. д' Орбиньи в 1842 г. Название
происходит от р. Об (латинское Alba), правого притока р. Сены.
Стратотипический разрез расположен юго-восточнее Парижа.
Альбский ярус охарактеризован большим числом аммонитов, на
основании которых проводится зональное деление. Как и во многих других случаях, стратотип альбского яруса не позволяет фаунистически обосновать ни нижнюю, ни верхнюю границы яруса и
дать зональное деление по аммонитам верхней части среднего и
всего верхнего альба из-за недостаточной их охарактеризованности
аммонитами.
Сеноманский ярус выделен в 1847 г. А. д' Орбиньи в департаменте Сарта Франции близ г. Ле-Ман (старинное название — Сеnomanum). Эти отложения вначале включались им в состав туронского яруса, но затем, убедившись в существенных различиях
в фауне аммонитов и рудистов, д' Орбиньи выделил их в самостоятельный ярус, который был охарактеризован более чем
800 видами. Сеноман в стратотипе представлен терригенными породами, образовавшимися в гидродинамически неспокойной среде,
вследствие чего на нескольких уровнях имеются следы подводных
перерывов. Сеноманский ярус охарактеризован аммонитами, белемнитами, устрицами, брахиоподами. В настоящее время принято трехчленное деление сеномана.
Название «турон» было предложено А. д'Орбиньи в 1842 г.
для карбонатных отложений, развитых в окрестностях г. Тур
(древнее название Turones). Здесь развиты мел, мергели и известняки, изредка переслаивающиеся с песчаниками. В разрезе множество перерывов. Отложения изобилуют остатками аммонитов,
иноцерамов, устриц, рудистов, гастропод, морских ежей, брахиопод, а также остракод, фораминифер и отпечатками растений.
Современное трехчленное деление турона сложилось за пределами
стратотипического района. Для России наибольшее значение имеет деление по иноцерамам, разработанное в различных регионах
центральной части Европы.
Коньякский ярус свое название получил от г. Коньяк, расположенного в западной части департамента Шаранта во Франции.
Здесь развиты карбонатно-терригенные породы, переполненные
остатками устриц, брахиопод, морских ежей, мшанок, аммонитов
и рудистов. Ныне, однако, стало ясно, что разрез в г. Коньяк
охватывает лишь небольшую часть коньякского яруса в современном понимании его объема, что требует обоснования нового стратотипа. Коньякский ярус разделяется на две аммонитовые зоны.
334
Принятое в центральной части Европы зональное деление коньяка
основано главным образом на иноцерамах.
Название сантонского яруса дано по г. Сент (Sant) в департаменте Приморская Шаранта во Франции. В 1857 г. Г. Кокан отнес
к сантону мягкий мел с кремнями и остатками губок, брахиопод,
морских ежей и двустворок.
Кампанский ярус свое название получил от гряды холмов
Гран-Шампань. Положение верхней границы в стратотипическом
районе неопределенное. Кампанский ярус расчленяется на два
подъяруса и четыре зоны, охарактеризованные главным образом
аммонитами и орбитоидами.
Название маастрихтскому ярусу дано по г. Маастрихт в южной части голландской провинции Лимбурт А. Дюмоном в 1849 г.;
здесь распространены мел и мелоподобные известняки с остатками аммонитов и белемнитов. Под маастрихтским ярусом ныне
понимают отложения, заключающие Hoploscaphites constrictus.
Различают нижний Маастрихт с Acanthoscaphites tridens и примитивными белемнеллами и верхний Маастрихт, который состоит из
зон Belemnitella junior и Belemnitella kazimiroviensis. Верхняя
граница маастрихтского яруса фиксируется по исчезновению аммонитов, белемнитов и многих других макрофоссилий, а также по»
резкому изменению комплексов планктонных фораминифер и нанопланктона.
Сводные разрезы меловой системы показаны на рис. 17.1.
ОРГАНИЧЕСКИЙ МИР
Меловой период завершает мезозойскую эру, и поэтому его органический мир несет все черты, характерные для переходного
этапа. Если в раннемеловую эпоху господствующее положение занимает мезофитная флора, то в позднемеловую эпоху все возрастающее значение приобретают кайнофитные элементы — покрытосеменные растения.
В морских бассейнах мелового периода важнейшими группами
являются головоногие, двустворчатые и брюхоногие моллюски,
морские ежи, брахиоподы, губки, мшанки, шестилучевые кораллы,
фораминиферы (рис. 17.2). На границе юры и мела происходит
новое значительное обновление аммонитов. Хотя многие юрские
аммоноидеи вымирают, но сохраняются представители Phylloceratidae, Lytoceratidae. Наряду с плоскоспиральной раковиной Polyptycites, Parahoplites, Acanthoceras, Neocomites, Simbirskites появляются ранее неизвестные роды с аномальными раковинами
Crioceras, Crioceratites, прямыми Baculites, Scaphites, Ancyloceras, улиткообразными Turrilites. Наряду с нормальной лопастной
линией появляются формы с упрощенной сутурой (Tissotia). Возникают и широко распространяются гигантские формы Pachydiscus, Ammonitoceras, отдельные экземпляры которых обладают раковиной до 2 м в поперечнике.
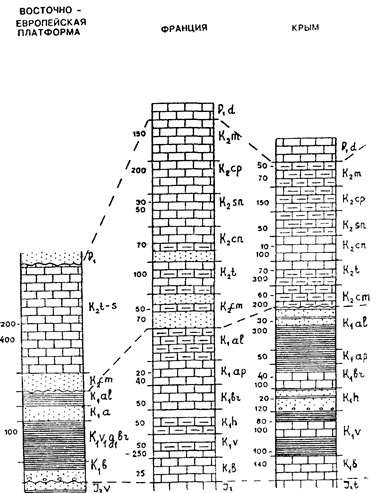
Рис. 17.1. Сопоставление сводных разрезов меловой системы. Условные
Значительно обновляется фауна белемнитов. В раннем мелу
наряду с Hibolites, Mesohibolites, Neohi'bolites, Cylindroteuthis,
Pachiteut'his встречаются своеобразные Duvalia. В позднемеловую
эпоху исключительно большим распространением пользуются
представители родов Actinocamax, Belemnitella, Belemnella. Переживают расцвет «неправильные» морские ежи и двустворчатые
моллюски. Среди последних большое стратиграфическое значение
имеют иноцерамы. Эти разнообразные по форме, очертаниям и
размерам моллюски распространены во всех морях. Достигают
расцвета устрицы родов Ostrea, Gryphaea, Exogyra и перешедшие
из позднеюрской эпохи бухии.
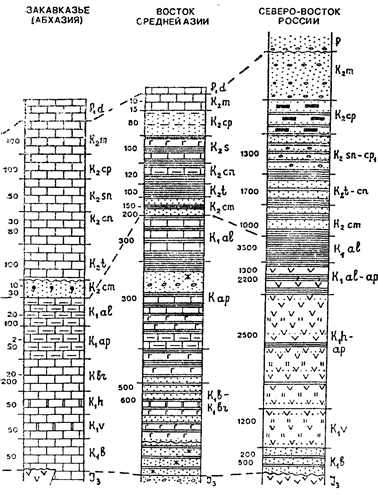
обозначения см. на рис. 8.1
В тропических морях широко развиты крупные толстостворчатые рудисты — Hippurites, Radiolites, Requenia, Toucasia, слагающие протяженные рифоподобные массивы. В их строении принимают участие брюхоногие моллюски — Nerinea в раннем мелу и
Acteonella — в позднем. Большую роль играют uyclostoma, Cyprea, Conus, 'Fusus, Murex. В составе гастропод большие изменения
Произошли в середине мелового периода, когда появилась основная масса семейств подотряда Neogastropoda, характерных для
кайнозойской эры.
Кораллы претерпели заметные изменения. Появились новые
семейства, рифообразующие формы. Ostreidae, Fungidae, Turbi-
22-1164 ' з37

Рис. 17.2. Характерные представители организмов мелового периода.
Аммоноидеи: / — Ancyloceras (Ki); 2 — Scaphites (Kz); 3 — Crioceras (Ki^;
4 — Schloenbachia (Kz); 5 — Simbirskites (Ki); 6 — Hoplites (Ki); 7—Tissotia
(Кг). Белемниты: 8 — Duvalia (Ki); 9 — Actinocamax (Kz); 10 — Belemnitella
(Кг)- Двустворчатые моллюски: 11 — Inoceramus retrorsus; 12 — Buchia (Aucella); 13 — Gryphaea; 14 — Hippurites. Морские ежи: 15 — Micraster; 16—
Echinocorys
nolidae нередку встречаются с рудистами. Характерной позднемеловой формой является одиночный коралл Cyclolites.
Брахиоподы, так обильно представленные в юрском периоде,
постепенно теряют свою ведущую роль. Много мшанок (около
1000 видов).
Во второй половине мелового периода произошла вспышка в
развитии губок. В позднемеловое время встречается большое количество Siphonia, Jerea, Ventricutites, Coeloptycium. В теплых
морях появились и широко распространились крупнораковинные
Orbitolina, в позднемеловое время — Alveolina, Orbitoides, а также ряд мелких форм: Globigerina, Miliolina, Textularia.
Среди неправильных морских ежей важное стратиграфическое
значение имеют в раннем мелу Toxaster, а в позднем — Micraster,
Holaster, Echinocorys. Из правильных морских ежей в меловом
периоде жили Cidaris, Acrocidaris, Salenia, а из криноидей бесстебельные Marsupites, Vintacrinus.
Среди костных рыб вначале преобладали лучеперые, но затем
они были вытеснены костистыми. С позднего мела началось развитие хрящевых рыб, в том числе высших акуловых.
Среди морских позвоночных продолжали существовать некогда
господствовавшие плезиозавры (рис. 17.3). Они дали начало причудливым и специализированным гигантам Elasmosaurus с необычайно длинной шеей. Ихтиозавры постепенно исчезают и уже в
конце раннемеловой эпохи вымирают. Им на смену приходит новая водная группа — змееподобные долихозавры и мозозавры,
которые быстро становятся господствующими в море.
Среди морских водорослей очень характерны микроскопические золотистые — кокколитофориды (нанопланктон) и диатомовые. Надо отметить, что нанопланктон и мелкие фораминиферы
в позднем мелу участвовали в формировании белого писчего мела.
Органический мир суши был своеобразен. В начале раннемеловой эпохи наземная флора имела много общего с позднеюрской.
Она состояла из цикадофитовых, гинкговых и папоротниковых, но
наряду с ними бурного расцвета достигли беннетиттовые. В целом.
флора раннего мела оставалась мезофитной, но в барреме появились первые покрытосеменные, которые стали преобладать в позднемеловое время, и флора приобрела облик кайнофитной. Если в;
барреме это были отдельные представители, то уже в альбе покрытосеменные встречаются очень часто. В сеном ане они оттесняют голосеменные. Кайнофитный облик позднемеловой флоры определяется развитием в ее составе большого количества таких современных форм, как дуб (Quercus), бук (Fagus), ива (Salix),.
береза (Betula), платан, лавр, магнолия.
В то время как в наземной флоре в меловом периоде произошли существенные изменения и рубежом может считаться альбский век, среди наземной фауны, особенно среди позвоночных, таких сильных изменений не наблюдается. Продолжали господствовать динозавры. Для раннего мела были характерны игуанодоны,
22* 33»
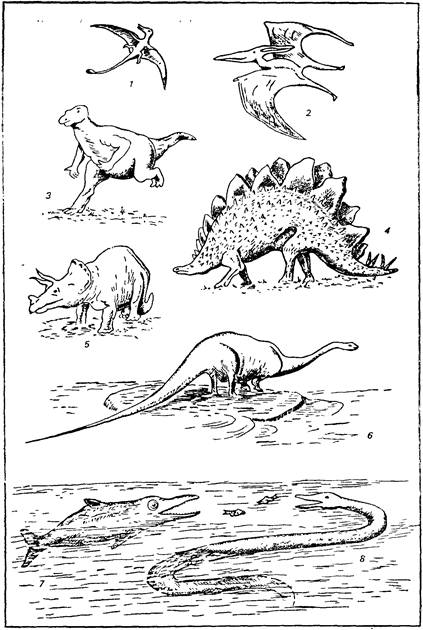
Рис. 17.3. Характерные представители мезозойских пресмыкающихся: 1 — Rhamphorynchus (Js); 2 — Pteranodon (Кг); 3 — Iguanodon (Ki); 4 — Stegosaurus
(Ja—Ki); 5 — Triceratops (Кг); 6 — Diplodocus (Js); 7 — Ichtyosaurus (Ji);
8 — Mososaurus (Кг)
для позднего — семейство гадрозавров или утконосых динозавров, которые были одними из самых крупных двуногих животных
того времени. Наряду с ними обитали рогатые травоядные четвероногие динозавры. Существовали и огромные хищные динозавры — тиранозавры, тарбозавры. Большим распространением
пользовались летающие ящеры — птеродактили. Все они доживут
до конца Маастрихта и затем навсегда исчезнут.
Для мелового периода характерно появление змей, которые
как и крокодилы, большое развитие получили в кайнозое
Млекопитающие, появившиеся еще в начале мезозоя все еще
были мелкими и встречались довольно редко, но за меловой период они прошли сложный эволюционный путь, дав начало многим
формам после исчезновения динозавров. В конце мелового периода появились мелкие сумчатые и эутерии.
В классе птиц в меловое время уже нет переходных форм.
Вместо них появились настоящие птицы — преимущественно зубастые, хотя известны и первые беззубые птицы. Зубастые птицы
вымерли в конце мелового периода. Особенно много насекомых
которые сильно эволюционировали в тесной связи с растениями
Начиная с сеноманского века состав насекомых стал приобретать
кайнозойский облик.
Дата добавления: 2016-04-06; просмотров: 957;