Соматовисцеральная система.
Соматовисцералъная система включает рецепторы трех типов по месту их расположения: кожные рецепторы, мышечные и суставные рецепторы — проприоцепторы и висцеральные рецепторы — инте-роцепторы. По функциям, независимо от места их расположения, рецепторы соматовисцеральной системы — это механо-, термо-, хеморецепторы, а также рецепторы боли — ноцицепторы. Общим свойством рецепторов соматовисцеральной системы является то, что они не образуют сенсорные органы, а широко распространены по всему телу. Кроме того, их афферентные волокна не образуют высокоспециализированные нервы, подобные зрительному и слуховому, а входят в многочисленные нервы тела и центральные тракты.
Кожная механорецепция.На различных участках кожи человека расположены инкапсулированные механорецепторы, которые иннер-вируются миелинизированными афферентными волокнами диаметром 5-12 мкм, проводящими импульсы со скоростью 30-70 м/с. Струк-
64

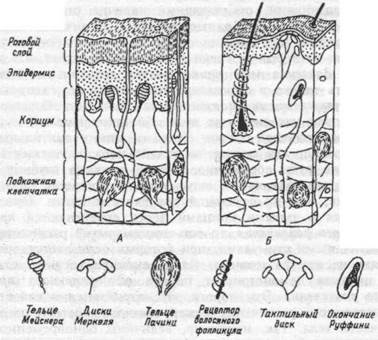
 турные особенности механорецепторов и их расположение на гладкой коже и на волосистых участках кожи иллюстрирует рис. 16.1. По функции все механорецепторы делятся на датчики давления стимула, датчики скорости его воздействия и датчики ускорения. Таблица 16.1 суммирует структурно-функциональные соотношения кожных рецепторов.
турные особенности механорецепторов и их расположение на гладкой коже и на волосистых участках кожи иллюстрирует рис. 16.1. По функции все механорецепторы делятся на датчики давления стимула, датчики скорости его воздействия и датчики ускорения. Таблица 16.1 суммирует структурно-функциональные соотношения кожных рецепторов.

|

 Рис.16.1. Механорецепторы кожи и их расположение на гладкой коже ладони (А) и волосистых участках кожного покрова (Б).
Рис.16.1. Механорецепторы кожи и их расположение на гладкой коже ладони (А) и волосистых участках кожного покрова (Б).
 Таблица 16.1.Классификация функциональных свойств кожных механорецепторов
Таблица 16.1.Классификация функциональных свойств кожных механорецепторов
 | |||
 |
| Медленная |
| Быстрая |
| Участки кожи не покрытые волосами |
| Волосистые участки кожи |
Адаптация к постоянному давлению
Тип рецепторов и их локализация
 Функциональное назначение
Функциональное назначение
Диск Мекреля
Тактильные диски, окончания Руффини
Датчик
интенсивности (рецепторы давления)
Тельце Мейснера
Рецептор
волосяного
фолликула
Датчики скорости
(рецепторы
прикосновения)
Очень быстрая
Тельце Пачини Тельце Пачини
Датчики
ускорения
(виброрецепторы)
 65
65


 В коже имеется также множество так называемых свободных нервных окончаний, на которых отсутствуют корпускулярные структуры. Афферентные волокна свободных нервных окончаний — не-миелинизированные аксоны со скоростью проведения 1 м/с. Геометрия свободных нервных окончаний весьма разнообразна. Свободные нервные окончания реагируют импульсом не более, чем на один уровень интенсивности — пороговый. Поэтому они считаются пороговыми датчиками, отмечающими наличие стимула в определенном месте кожи. Свободные нервные окончания принимают участие в передаче информации о слабых (околопороговых) движущихся по коже стимулах и имеют отношение к ощущению щекотки.
В коже имеется также множество так называемых свободных нервных окончаний, на которых отсутствуют корпускулярные структуры. Афферентные волокна свободных нервных окончаний — не-миелинизированные аксоны со скоростью проведения 1 м/с. Геометрия свободных нервных окончаний весьма разнообразна. Свободные нервные окончания реагируют импульсом не более, чем на один уровень интенсивности — пороговый. Поэтому они считаются пороговыми датчиками, отмечающими наличие стимула в определенном месте кожи. Свободные нервные окончания принимают участие в передаче информации о слабых (околопороговых) движущихся по коже стимулах и имеют отношение к ощущению щекотки.
Ощущение давления или прикосновения (тактильные ощущения) можно вызвать только в определенных точках кожи, которые называются чувствительными тактильными точками. Большое число тактильных точек находится на коже кончиков пальцев и на губах. На коже, покрывающей плечи, бедра и спину тактильные точки встречаются редко, для появления тактильного ощущения на внутренней стороне руки достаточно вдавить кожу на глубину 0,01 мм (10 мкм). Пороги тактильных ощущений на кончиках пальцев значительно ниже, чем на остальной поверхности кисти. При помощи ножек циркуля с тупыми концами легко определяются пространственные пороги различения, то есть минимальные расстояния между двумя тактильными стимулами, при которых оказывается возможным воспринять их как раздельные. Одновременное касание кожи кончиками циркуля демонстрирует пороги одновременного пространственного различения. Эти пороги рассматриваются в качестве меры пространственного различения кожей тактильных стимулов в исследуемой части тела. Так, например, величины одновременного пространственного различения кончика языка, пальцев и губ — порядка 1-3 мм — свидетельствуют о высокой чувствительности этих областей тела. На спине, плечах и бедрах пространственное различение значительно хуже — 50- 100 мм. Различия чувствительности связаны с плотностью рецепторов в тех или иных областях тела.
Из всех кожных механорецепторов наиболее изучены тельца Па-чини — детекторы коротких механических воздействий. Многократное частое раздражение этих рецепторов вызывает ощущение вибрации. Капсула тельца Пачини состоит их множества одинаковых слоев, вложенных друг в друга наподобие "матрешки". В сердцевине "матрешки" находится нервное окончание. Эта многослойная конструкция работает как высокочастотный фильтр: при кратковременном воздействии капсула деформируется и действует на нервное окончание. Если действие механического раздражителя продолжается, то происходит изменение расстояний между слоями капсулы, которое компенсирует постоянное давление на тельце Пачини и приводит к отсутствию деформации нервного волокна.
Капсула тельца Пачини деформируется при воздействии механического раздражителя и действует на нервное окончание. В нервном окончании возникает генераторный потенциал. Поскольку капсула работает как высокочастотный фильтр, то она не реагирует на сти-
66

 муляцию постоянным давлением. В самом возбудительном процессе нейрона капсула не участвует. Она только меняет характер раздражения. При многократном раздражении генераторные потенциалы телец Пачини снижаются.
муляцию постоянным давлением. В самом возбудительном процессе нейрона капсула не участвует. Она только меняет характер раздражения. При многократном раздражении генераторные потенциалы телец Пачини снижаются.
Потенциалы действия нервных волокон, отходящих от телец Пачини. следуют частоте раздражения до 200-400 Гц, в интервале 400- 500 Гц соответствие импульсации частоте стимула нарушается, выше 500 Гц они перестают реагировать. Люди при одинаковом раздражении кожи ладони отмечают два вида ошушений — вибрации и трепетания. Для возникновения ощущения вибрации необходима более высокая частота и меньшая амплитуда, чем для ощущения трепетания, которое появляется при более низких частотах, но большей амплитуде колебаний. Трепетание исчезает при поверхностной анестезии, так же как разряды рецепторов, реагирующих на низкие частоты вибрации. Местная анестезия поверхностных слоев кожи не уничтожает вибрационную чувствительность и ответы высокочастотных рецепторов. Условием появления ощущения вибрации в коже является одновременное вовлечение в реакцию нескольких телец Пачини, поскольку именно в этом случае разряд в афферентных волокнах становится эффективным стимулом.
Терморецепция.Терморецепция или температурное чувство соотносится с двумя ощущениями — тепла и холода. Эти ощущения можно вызвать в температурных точках тела. Специальные рецепторы тепла и холода выполняют две основные функции — они отвечают на изменения температуры окружающей среды и участвуют в регуляции температуры тела (рис. 16.2).
 | |||
 | |||
|
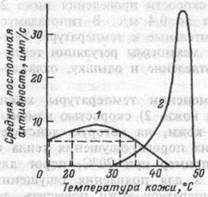
Рис. 16.2. Функция температурных рецепторов кожи (схема).
1 — хоподовые рецепторы —
частота разряда мало зависит
от температуры;
2 — тепловые рецепторы —
выраженная реакция возникает
в узком диапазоне темпера
тур.


 Рецепторы кожи, чувствительные к холоду и теплу (точки тепла и холода), расположены на разных участках тела. Плотность и общее число температурных точек меньше, чем число тактильных точек во всех областях тела человека. Точек холода на коже значительно больше, чем точек тепла. Так, например, на поверхности кисти руки имеется по 1-5 точек холода на каждом квадратном сантиметре и лишь в среднем по 0,4 точки тепла. Максимальная плотность тех и
Рецепторы кожи, чувствительные к холоду и теплу (точки тепла и холода), расположены на разных участках тела. Плотность и общее число температурных точек меньше, чем число тактильных точек во всех областях тела человека. Точек холода на коже значительно больше, чем точек тепла. Так, например, на поверхности кисти руки имеется по 1-5 точек холода на каждом квадратном сантиметре и лишь в среднем по 0,4 точки тепла. Максимальная плотность тех и
«57




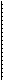 других точек характерна для кожи лица, наиболее чувствительной к изменениям температуры. Число точек холода здесь достигает 16- 19 на см2.
других точек характерна для кожи лица, наиболее чувствительной к изменениям температуры. Число точек холода здесь достигает 16- 19 на см2.
Низкая, по сравнению с плотностью механорецепторов, плотность точек холода и, особенно, тепла свидетельствует, что одновременные пространственные пороги для температурных стимулов относительно велики. Пространственные пороги для холодовых стимулов ниже, чем для тепловых. Так, на бедре одновременный пространственный порог для тепловых стимулов в продольном направлении равняется 26 см, а в поперечном — 9 см. Холодовым стимулам соответствуют значения 16,5 см и 2,9 см.
Специфические терморецепторы (рецепторы холода и тепла) предположительно представляют собой свободные нервные окончания, которые несут также механорецепторные функции. У человека рецепторы холода располагаются в эпидермисе и непосредственно под ним, а рецепторы тепла — преимущественно в верхнем и среднем слоях собственно кожи. Рецепторы холода связаны с тонкими миелинизи-рованными волокнами, а рецепторы тепла — с немиелинизированны-ми волокнами. Специфические терморецепторы характеризуются рядом общих показателей. Так, при постоянной температуре кожи они тонически разряжаются с частотой, зависящей от температуры — статическая реакция. В случае изменения температуры кожи они увеличивают или уменьшают частоту разряда — динамическая реакция. Специфические терморецепторы нечувствительны к нетепловым стимулам, а пороги их импульсной реакции сравнимы с порогами появления ощущений при тепловой стимуляции кожи. Они обладают малыми рецептивными полями (1 мм2 и менее), и каждое афферентное волокно обслуживает только одну точку тепла или холода. Кроме того, у этих афферентных нервных волокон скорости проведения ниже 20 м/с, а у некоторых видов они доходят до 0,4 м/с. В гипоталамусе головного мозга есть рецепторы, чувствительные к температуре. Температура этой области мозга активирует механизмы регуляции температуры тела: нагревание вызывает потоотделение и одышку, охлаждение — сужение сосудов и дрожь.
Ощущения, появляющиеся при изменении температуры кожи, определяются: 1)исходной температурой кожи, 2) скоростью изменения температуры кожи, 3) площадью кожи, на которую действует стимул. Исходная температура влияет на пороги ощущения тепла и холода. При низких температурах, например при 20°С, порог для появления ощущения тепла высокий, а для появления ощущения холода — низкий. Если исходную температуру кожи повысить, то тепловые пороги уменьшатся, а холодовые увеличатся. При этом следует иметь в виду, что в зависимости от условий стимуляции одна и та же температура кожи может вызывать и ощущение тепла, и ощущение холода. Если, например, исходная температура равна 32°С, то нагревание на 0,5°С вызовет ощущение тепла, а при исходной температуре равной 33°С, охлаждение на 0,5°С — отчетливое ощущение холода. Однако, в том и другом случае конечная температура кожи будет одинаковой — 32,5°С.
68



 Скорость изменения температуры, если она превышает 0,1°С с (6°С мин), почти не оказывает влияния на пороги ощущения тепла и холода. При снижении скорости изменения температуры оба порога монотонно возрастают.
Скорость изменения температуры, если она превышает 0,1°С с (6°С мин), почти не оказывает влияния на пороги ощущения тепла и холода. При снижении скорости изменения температуры оба порога монотонно возрастают.
Площадь кожи, на которой меняется температура, отражается на порогах ощущений при охлаждении и нагревании, соответственно, больше в случае малых площадей, чем при больших. При подобном надпороговым изменении температуры кожи интенсивность ощущений возрастает с увеличением стимулируемой площади. Следовательно, и в околопороговом, и в надпороговом диапазонах происходит пространственная суммация импульсных реакций терморецепторов в нервных центрах.
Проприоцепция.Проприоцепцией называется совокупность способностей человека ориентироваться в положении своих конечностей по их отношению друг к другу, воспринимать собственные движения и оценивать сопротивление совершаемым движениям. Источником эффективных стимулов во всех этих случаях является само тело, в мышцах, сухожилиях и суставах которого преимущественно расположены соответствующие рецепторы — проприоцепторы.
Проприоцепция, как модальность, обладает тремя качествами. Это, во-первых, ощущение положения конечностей, базирующееся на информации об углах в каждом суставе. Так, человек может судить об относительном положении конечностей. Поскольку ощущение положения конечностей хорошо сохраняется, например, после долгого сна, оно лишь в незначительной степени подвержено адаптации. Во-вторых, проприоцепция обеспечивает ощущение движения. В этом случае проприоцепторы воспринимают и направление, и скорость движения при изменениях суставного угла даже без зрительного контроля (например, сгибание и разгибание руки в локте).
Порог восприятия движения зависит от величины и скорости изменения угла. Для проксимальных суставов (например, плечевых) — он ниже, чем для дистальных (например, межфаланговых суставов пальцев).
Третьим качеством проприоцепции является ощущение усилия. Проприоцепторы способны оценивать величину мышечного усилия, необходимого для совершения определенного движения, а также для сохранения некоторого положения сустава при действии того или иного сопротивления движению.
Проприоцепторы находятся в подкожных структурах — мышцах, сухожилиях и суставных сумках. В мышцах — это мышечные веретена, а в сухожилиях — сухожильные органы Гольджи. Они участвуют в восприятии положения и движения тела и конечностей. В суставных сумках это рецепторы типа окончаний Руффини, сухожильных органов Гольджи и, в меньшем числе, рецепторы типа телец Пачини. Эти рецепторы связаны с миелинизированными нервными волокнами. Кроме того, в суставных сумках имеются свободные нервные окончания с немиелинизированными афферентами. Суставные механорецепторы при изменениях положения сустава и
69

 соответствующих сжатиях или растягиваниях суставной сумки, передают информацию о положении суставов, направлении и скорости движения конечностей тела. Фазно-тонические свойства суставных рецепторов проявляются не только тогда, когда сустав движется так, что обусловливает возрастание активности рецептора, но и тогда, когда движение осуществляется в обратном направлении. Работу рецепторов положения суставов объясняет рис. 16.3.
соответствующих сжатиях или растягиваниях суставной сумки, передают информацию о положении суставов, направлении и скорости движения конечностей тела. Фазно-тонические свойства суставных рецепторов проявляются не только тогда, когда сустав движется так, что обусловливает возрастание активности рецептора, но и тогда, когда движение осуществляется в обратном направлении. Работу рецепторов положения суставов объясняет рис. 16.3.
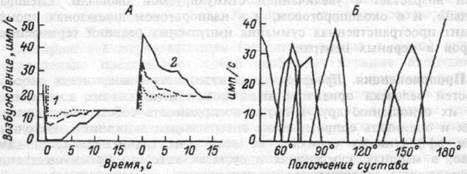
Рис.16.3. Функция рецепторов положения суставов.
На А — реакция неполностью адаптирующегося рецептора
коленного сустава сгибанием (I) и разгибанием (2).
Сплошные линии — быстрое сгибание и разгибание, штриховые
и пунктирные линии — более медленное.
На Б — области ответа или "рецептивные углы" нескольких
суставных рецепторов при их постоянном возбуждении в
результате длительного расположения сустава под разными
углами.
 Общее представление об относительном положении суставов формируется у человека в результате процесса интеграции информации от рецепторов различных мышц, сухожилий и суставов. Этот процесс начинается, как и в других сенсорных системах, на уровне подкорковых ядер.
Общее представление об относительном положении суставов формируется у человека в результате процесса интеграции информации от рецепторов различных мышц, сухожилий и суставов. Этот процесс начинается, как и в других сенсорных системах, на уровне подкорковых ядер.
В центральной интеграции, кроме того, используются и сигналы несенсорного происхождения. Речь идет, в частности, об эфферентных "копиях приказов мышцам", которые центральные отделы моторной системы адресуют в центральную нервную систему для взаимодействия с сенсорными сигналами, поступающими от пропри-оцепторов. Эти эфферентные "копии" содержат информацию о планируемой активности мышц и о том движении, которое последует. Подобные сведения важны для выделения из афферентной информации о движении той афферентации, которая связана с запрограммированной мышечной активностью.
Интероцепция.Интероцепцией называют восприятие раздражений (стимулов), поступающих из внутренней среды организма: от орга-
70

 нов и тканей. Многочисленные интероцепторы, расположенные в стенках сосудов и в различных органах и тканях с учетом их фун-кионального назначения имеханизма возбуждения, подразделяют на механорецепторы, хеморецепторы, осморецепторы и терморецепторы.
нов и тканей. Многочисленные интероцепторы, расположенные в стенках сосудов и в различных органах и тканях с учетом их фун-кионального назначения имеханизма возбуждения, подразделяют на механорецепторы, хеморецепторы, осморецепторы и терморецепторы.
К механорецепторам относят рецепторы кровеносных и лимфатических сосудов, рецепторы сердца, легких и других структур, способных под влиянием механического раздражения растягиваться и деформироваться. К этой группе интероцепторов относят также проприоцепторы мышц и сухожилий.
К не менее обширной группе интероцепторов — хеморецепторам — причисляется вся масса тканевых рецепторов, реагирующих на различные химические вещества. Это рецепторы аортального и ка-ротидного клубочков, рецепторы слизистых оболочек пищеварительного тракта и органов дыхания, рецепторы серозных оболочек. К группе хеморецепторов относятся осморецепторы, выделяемые среди интероцепторов на основе их избирательной чувствительности к изменениям концентрации осмотически активных веществ в крови и внеклеточной жидкости. Следует также иметь в виду, что термином хеморецепторы обозначаются рецепторы вкуса и обоняния, стимулируемые из внешней среды (экстероцепция), которые будут рассмотрены отдельно.
Существование термочувствительных интероцепторов — терморецепторов — подтверждено только для верхних отделов пищеварительного тракта. Об их наличии в других внутренних органах судят по косвенным данным. Интероцепторы представлены в организме свободными нервными окончаниями, а также различными видами сложных инкапсулированных рецепторов, например, телец Пачини.
Ноцицепция.Ноцицептивная чувствительность — это восприятие стимулов, вызывающих в организме ощущение боли. Ощущение боли коренным. образом отличается от других сенсорных модальностей содержанием информации, которую она в себе заключает. Боль информирует организм об опасности, о нарушениях в процессах, протекающих в различных органах и тканях, поскольку вызывается повреждающими, вредными для организма стимулами. Трудно переоценить значение этой информации для клиники, так как именно болевое проявление действия вредных факторов чаще всего приводит пациента к врачу. Более того, ощущение боли, при всей его субъективности, оказывается часто решающим для диагноза заболевания.
Качества боли классифицируются с учетом места их происхождения и свойств (рис. 16.4). Модальность "боль" сочетает два типа соматических болей, а также висцеральные боли. Поверхностной болью называется соматическая боль кожного происхождения, а если ее источник локализован в мышцах, костях, суставах и соединительной ткани, она называется глубокой болью. Глубокая боль, как правило, тупая, плохо локализуется, обладает тенденцией ирради-ировать в окружающие структуры, сопровождается неприятными ощущениями, тошнотой, сильным потоотделением, падением арте-
71




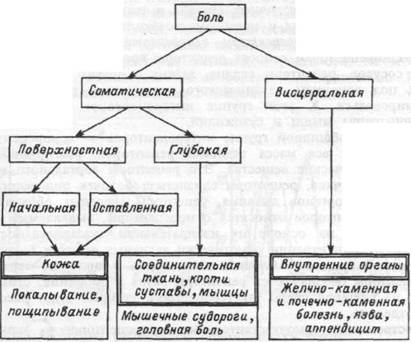
Рис.16.4. Классификация видов боли и их локализация.
 риального давления. Аналогичная картина характерна и для висцеральных болей.
риального давления. Аналогичная картина характерна и для висцеральных болей.
Укол иглой, провоцирующий поверхностную боль, вызывает легко локализуемое ощущение, быстро исчезающее с прекращением стимуляции. Эта начальная (первая) боль, с латентным периодом до 0,2 с, при высоких интенсивностях стимуляции часто сопровождается отставленной (второй) болью, имеющей уже не острый, а тупой или ноющий характер, с латентным периодом 0,5-1 с. Отставленная боль пространственно ощущается как более диффузная, исчезает медленно, иногда ей сопутствует тошнота, другие негативные реакции организма.
Расположенные в коже специальные болевые рецепторы — ноци-цепторы — подразделяются на чисто механочувствительные и меха-нотермочувствительные. Последние называются полимодальными ноцицепторами и их число в коже человека превышает количество ноцицепторов других типов. Ноцицепторы кожи, также как и внутренних органов и скелетных мышц, представляют собой свободные нервные окончания с тонкими миелинизированными (скорость проведения приблизительно 11 м/с) или немиелинизированными (скорость проведения окло 1 м/с) нервными волокнами. При поверхностной боли сигналы начальной боли передаются по тонким ми-
72


 елинизированным волокнам, а сигналы отставленной боли — по немелинизированным волокнам. Это является свидетельством того, что различие во времени появления ощушений начальной и отставленной боли связаны, главным образом, с разными скоростями проведения по соответствующим нервным волокнам.
елинизированным волокнам, а сигналы отставленной боли — по немелинизированным волокнам. Это является свидетельством того, что различие во времени появления ощушений начальной и отставленной боли связаны, главным образом, с разными скоростями проведения по соответствующим нервным волокнам.
В полых внутренних органах, образованных гладкими мышцами, находятся механочувствительные висцеральные ноцицепторы, часть которых реагирует на пассивное растяжение, а другая часть на активное напряжение гладких мышц. Множество ноцицепторов, активируемых раздражающими газами или частицами пыли, содержат легкие. Однако, пока еще не известно, чем вызываются ощущения сильных висцеральных болей — сопутствующими механическими явлениями или химическими изменениями в тканях. Предполагается, что по характеру возникновения возбуждения ноцицепторы относятся к хеморецепторам. Химическими раздражителями являются вещества, которые до раздражения находятся внутри клеток (брадикини-ны, ионы калия).
Существуют особые виды боли — проецируемая и отраженная. В качестве примера причины проецируемой боли можно привести резкий удар по локтевому нерву, проходящему в области локтя у самой поверхности. Подобный удар вызывает очень неприятное, трудноописываемое ощущение, распространяющееся на те участи руки, которые обслуживаются этим нервом — на участок от локтя до кисти и на саму кисть. В принципе, проецируемые ощущения могут возникать в рамках любой сенсорной модальности, причем, многие длительные проецируемые боли, в отличие от боли от удара по локтевому нерву, имеют клиническое значение. Механизм их возникновения объясняется следующим. Сигналы, вызываемые ненормальной стимуляцией афферентного волокна, передаются обычно в мозг по латеральному спинно-таламическому тракту, порождая там ощущения, относимые к области, которую иннервирует данное афферентное волокно. Одна из распространенных причин проецируемых болей — это пережатие спинальных нервов в местах их вхождения в спинной мозг в результате повреждения межпозвонковых хрящевых дисков. Афферентные импульсы в ноцицептивных волокнах при такой патологии вызывают болевые ощущения, которые проецируются в область, связанную с травмируемым спинальным нервом.
Отраженная боль отличается от проецирумой боли тем, что она вызывается не прямой стимуляцией нервных волокон, а раздражением каких-либо рецептивных окончаний. Отраженными болями называются болевые ощущения не во внутренних органах, от которых поступают болевые сигналы, а в расположенных на некотором расстоянии от больного органа поверхностных областях. Хорошо известный пример отраженной боли — боль, возникающая в сердце, но ощущаемая в плече и в узкой полоске на медиальной поверхности руки. Так как взаимоотношения между отдельными участками кожи (дерматомами) и внутренними органами хорошо известны, подобные отраженные боли играют большую роль в диагностике
73


 различных заболеваний. Механизм возникновения отраженной боли сводится к следующему. Некоторые кожные болевые афференты и болевые афференты, идущие от внутренних органов, при вхождении в один и тот же сегмент спинного мозга могут конвергировать на один и тот же нейрон. Возбуждение таких клеток на периферии интерпретируется человеком как боль.
различных заболеваний. Механизм возникновения отраженной боли сводится к следующему. Некоторые кожные болевые афференты и болевые афференты, идущие от внутренних органов, при вхождении в один и тот же сегмент спинного мозга могут конвергировать на один и тот же нейрон. Возбуждение таких клеток на периферии интерпретируется человеком как боль.
Примером проявления конвергенции ноцицептивных афферентов отдерматомов и от внутренних органов на одни и те же нейроны, является гиперчувствительность кожи — гиперпатия — в соответствующих дерматомах. Она возникает в результате повышения возбудимости вставочных нейронов под действием висцеральных импульсов. Известно явление "солнечного ожога", возникающее после повреждения кожи интенсивным ультрафиолетовым облучением и проявляющееся покраснением кожи, повышением ее чувствительности кмеханическим стимулам. Аналогичные явления сопровождают повреждения кожи нагреванием, охлаждением, рентгеновскими лучами и механической травмой. Понижение болевого порога называется гипералгезией. Повышение болевого порога — гипалгезия, равно как и полная потеря болевой чувствительности — аналгезия, устанавливаются обычно лишь в сочетании с нарушением или ослаблением других модальностей кожной рецепции.
Субъективные ощущения боли зависят не столько от интенсивности стимула, сколько от состояния эмоциональной, мотивацион-ной и волевой сферы человека. Уменьшению боли способствует отвлечение внимания, стресс, гипнотическое внушение, экстремальные ситуации и др. Адаптация к боли в физиологическим смысле не существует: болевые пороги при повторном раздражении не изменяются или даже снижаются, то есть наблюдается сенсибилизация (повышение чувствительности) участка кожи, стимулируемого болевым раздражителем. Такой сенсетизации способствует включение эмоционального компонента реакции на раздражитель приизмерениях порогов боли.
Центральные проекции соматовисцеральной системыпредставлены на рис. 16.5. Чувствительные нервные волокна в составе спи-нальных нервов идут к спинному мозгу и несут информацию от всех рецепторов — тактильных, температурных, болевых. Афферен-тация, связанная с тонкими механическими ощущениями — вибрацией, оценкой положения и движений суставов, осязанием мелких шероховатостей поверхностей, т.е. ощущений с высоким пространственным разрешением — проводится по волокнам задних столбов (пучкиГолля и Бурдаха). Эти волокна первого порядка заканчиваются в ядрах задних столбов — под самым основанием черепа. Отсюда после переключения отходит новая группа аксонов, образующих медиальную петлю, которая после пересечения средней линии в области ствола мозга заканчивается в вентро- базальной группе ядер таламуса. Здесь начинается новая группа аксонов, которые заканчиваются в первичной соматической проекционной зоне коры головного мозга — S, (поля 1,2,3 по Бродману). Афферентация, связанная с ощущениями плохо локализуемого давления, температуры и

 боли в составе передне-бокового пучка восходит по спино-талами-ческому спино-ретикуло-таламическому путям, которые содержат нейроны в основном третьего-четвертого порядков к неспецифическим ядрам таламуса, откуда широко проецируются в разные области коры. Это мультисинаптическая афферентная система, в которой имеются короткоаксонные вставочные клетки на всем ее протяжении, причем на вставочных нейронах спинного мозга заканчиваются первичные афференты от рецепторов.
боли в составе передне-бокового пучка восходит по спино-талами-ческому спино-ретикуло-таламическому путям, которые содержат нейроны в основном третьего-четвертого порядков к неспецифическим ядрам таламуса, откуда широко проецируются в разные области коры. Это мультисинаптическая афферентная система, в которой имеются короткоаксонные вставочные клетки на всем ее протяжении, причем на вставочных нейронах спинного мозга заканчиваются первичные афференты от рецепторов.

Рис.16.5. Схема организации соматосенсорной системы —
специфической (А) и неспецифической (Б) частей.



 Разные тракты, составляющие соматические афферентные пути действуют как единая система, а не как отдельные независимые каналы связи. Так, считают, поскольку часть волокон спино-тала-мического тракта заканчивается не только в неспецифических ядрах таламуса, но и в вентробазальной группе таламических ядер, где смешиваются с окончаниями волокон медиальной петли. Данные о ходе путей от рецепторов к мозгу соотносятся с концепцией об эпикритической (точной) и протопатической (диффузной) чувствительности.
Разные тракты, составляющие соматические афферентные пути действуют как единая система, а не как отдельные независимые каналы связи. Так, считают, поскольку часть волокон спино-тала-мического тракта заканчивается не только в неспецифических ядрах таламуса, но и в вентробазальной группе таламических ядер, где смешиваются с окончаниями волокон медиальной петли. Данные о ходе путей от рецепторов к мозгу соотносятся с концепцией об эпикритической (точной) и протопатической (диффузной) чувствительности.
Свойства нейронов лемнисковой системы следующие. Они, во-первых, имеют небольшие рецептивные поля, связанные с ограниченными, малыми участками кожной поверхности. Рецептивные поля центральных нейронов лемнисковой системы расширяются в каждом следующем синаптическом переключении. Во-вторых, лемнисковые нейроны сохраняют специфическую модальность. Например, нейрон, который возбуждается при воздействии механического стимула, не чувствителен к температурному воздействию или к движению сустава. В-третьих,функция зависимости числа импульсов от силы раздражения — линейная или степенная — всегда зависит от функции кожного рецептора, связанного с лемнисковым нейроном.
Нейроны спино-таламической системы имеют по сравнению с лемнисковыми более широкие рецептивные поля, отвечают на разные стимулы, т.е. являются мультимодальными. Функция зависимости частоты импульсации от силы стимула не связана с аналогичной характеристикой какого-либо из рецепторов.
Топографическое представительство периферических рецепторов, или карты, создаваемые проекцией афферентных волокон на ядрах мозгового ствола, промежуточного мозга или коры не отражают реальных проекций тела в объеме мозга. Особое значение в величине проекции играет плотность иннервации кожи, поэтому участки тела с наиболее обильной рецепцией имеют большой объем проекций в центрах мозга: например, рука, рот, язык.
Различия в структуре импульсной активности в зависимости от природы стимула четко проявляются в нейронах ядер задних столбов. В афферентах первого порядка этих отличий еще нет. Например, нейроны, которые активируются волокнами, идущими от рецепторов волосяных фолликулов, всегда отвечают импульсами, сгруппированными по два-три, следующими друг за другом. Клетки, активируемые волокнами, сигнализирующими о движении или положении суставов, дают длительные потоки импульсов, а не сгруппированные в пачки. Нейроны, реагирующие на постоянное давление на кожу, отвечают пачками импульсов. Эти данные говорят о кодировании "качеств" стимулов порядком следования нервных импульсов, их узором.
В вентробазальной области таламуса, где заканчивается медиальная петля, нейроны высокоспециализированы, модально обособлены (аналогично ядрам задним столбов спинного мозга). Здесь четко представлены данные о положении суставов. Таламические нейроны, связанные с проприоцепторами, реагируют на половину объема дви-
76



 жения в суставе. Например, для коленного или локтевого суставов половина клеток возбуждается при сгибании, половина — при разгибании, причем, максимальная импульсация возникает на одно из крайних положений — на полное сгибание или на полное разгибание. Для таких нейронов эквивалентом "интенсивности" стимула является положение сустава. Топография проприоцептивных талами-ческих нейронов сигнализирует об анатомическом положении сустава, и, таким образом, эти нейроны идентифицируют действующий сустав, а частота разряда в них показывает, под каким углом сустав согнут.
жения в суставе. Например, для коленного или локтевого суставов половина клеток возбуждается при сгибании, половина — при разгибании, причем, максимальная импульсация возникает на одно из крайних положений — на полное сгибание или на полное разгибание. Для таких нейронов эквивалентом "интенсивности" стимула является положение сустава. Топография проприоцептивных талами-ческих нейронов сигнализирует об анатомическом положении сустава, и, таким образом, эти нейроны идентифицируют действующий сустав, а частота разряда в них показывает, под каким углом сустав согнут.
Нейроны соматической проекционной коры делят на несколько групп: 1) реагирующие на движение волос; 2) быстроадаптирующие-ся клетки, отвечающие на раздражение поверхности кожи; 3) быстро адаптирующиеся клетки, возбуждаемые действием раздражителя на более глубокие ткани кожи - на тельца Пачини; 4) медленно адаптирующиеся клетки, отвечающие на стационарную стимуляцию — на диски Меркеля; 5) нейроны, связанные с проприоцепторами глубоких тканей, сигнализирующих о движении и положении суставов; 6) нейроны, реагирующие на вибрацию.
В трактовке способов кодирования соматосенсорной чувствительности много трудностей, связанных с недостаточностью и противоречивостью фактов, полученных преимущественно на животных.
Висцеральные рецепторы связаны с мозгом с помощью волокон черепно-мозговых нервов. В основном, это аксоны с медленной скоростью проведения, проходящие в задних и вентролатеральных столбах спинного мозга. Они заканчиваются в различных ядрах ствола мозга и таламуса (задневентральное и хвостатое ядро), причем, в таламических ядрах конвергируют соматическая, проприоцептивная и интероцептивная афферентация. Особенно важная роль в интеро-цептивной интеграции принадлежит гипоталамусу и разным частям лимбической системы мозга. Проекции висцеральных нервов обнаружены также в мозжечке.
В различных областях соматосенсорной и двигательной коры имеются представительства различных внутренних органов. Однако функциональная роль высших корковых проекций, равно как и входящих в их состав нейронов, не определена. И.М.Сеченов придавал большое значение "темным ощущениям", возникающим в результате раздражения рецепторов внутренних органов. Эти ощущения, наряду с экстроцептивными, формируют поведение человека, хотя в норме находятся по интенсивности на околопороговых уровнях, увеличиваясь при различных нарушениях внутренней среды организма.
Болевые ощущения передаются в мозг с помощью двух систем нейронов — медиальной и латеральной. Медиальная система проходит через центральные участки ствола мозга. Она ответственна за стойкую (тоническую) боль, передает сигналы в лимбическую систему головного мозга, участвующую в эмоциональном поведении. Именно этой медиальной системе приписывают эмоциональный компонент боли, который отражается в таких ее характеристиках,

 как "ужасная", "жестокая", "невыносимая", "душераздирающая" и т.п. Медиальная система боли состоит преимущественно из мелких волокон и завершается в таламической области мозга. Эта система проводит сигналы медленно, не приспособлена для точного и быстрого проведения информации о сильных раздражителях в критических ситуациях. Она передает диффузные неприятные ощущения. Другая — латеральная система боли — состоит из нервных трактов, проецирующихся в соматосенсорную кору головного мозга. Она наиболее активна при внезапной и резкой (фазической) боли, боли с четко выраженной локализацией. Латеральные пути отвечают за сенсорное качество боли, т.е. за характер ощущения — пульсирующая боль, укол, жжение и т.д. Активность латеральной системы быстро затухает, поэтому фазическая боль кратковременна, она подвергается мощному торможению со стороны других структур. Например, торможение исходит из центрального околоводопроводного серого вещества среднего мозга: по нисходящему тормозному тракту в задние рога спинного мозга, куда входят нервы от периферических рецепторов. Нисходящая тормозная система влияет на латеральную систему боли сильнее, чем на медиальную; при сильных болевых раздражителях эта нисходящая система активизируется эндогенными, т.е. образующимися в самом организме, опиоидами — эндор-финами и энкефалинами.
как "ужасная", "жестокая", "невыносимая", "душераздирающая" и т.п. Медиальная система боли состоит преимущественно из мелких волокон и завершается в таламической области мозга. Эта система проводит сигналы медленно, не приспособлена для точного и быстрого проведения информации о сильных раздражителях в критических ситуациях. Она передает диффузные неприятные ощущения. Другая — латеральная система боли — состоит из нервных трактов, проецирующихся в соматосенсорную кору головного мозга. Она наиболее активна при внезапной и резкой (фазической) боли, боли с четко выраженной локализацией. Латеральные пути отвечают за сенсорное качество боли, т.е. за характер ощущения — пульсирующая боль, укол, жжение и т.д. Активность латеральной системы быстро затухает, поэтому фазическая боль кратковременна, она подвергается мощному торможению со стороны других структур. Например, торможение исходит из центрального околоводопроводного серого вещества среднего мозга: по нисходящему тормозному тракту в задние рога спинного мозга, куда входят нервы от периферических рецепторов. Нисходящая тормозная система влияет на латеральную систему боли сильнее, чем на медиальную; при сильных болевых раздражителях эта нисходящая система активизируется эндогенными, т.е. образующимися в самом организме, опиоидами — эндор-финами и энкефалинами.
В результате нарушения процесса переработки болевых сигналов в центральной нервной системе, чувствительность, как правило, не исчезает — видоизменяются болевые ощущения. Нормальные болевые ощущения возможны только в условиях "правильной" активации корковых и подкорковых отделов центральной нервной системы, относящихся к ноцицептивной чувствительности. Так, если повреждение затрагивает таламические структуры, участвующие в проведении болевых сигналов, болевые стимулы вызывают чрезвычайно неприятные ощущения. Возникают также невыносимые спонтанные боли (таламические боли) в контралатеральной половине тела, включается также эмоционально- мотивационный компонент боли.
16.3. Слуховая система.
Слуховая система (слуховой анализатор) представляет собой совокупность механических, рецепторных и нервных структур, воспринимающих и анализирующих звуковые колебания. Звук — это колебательные движения упругих тел, распространяющиеся в различных средах в виде волн.
Звуковые волны обладают двумя переменными параметрами: частотой и амплитудой. Скорость распространения звука — величина постоянная и зависит только от среды, в которой он распространяется. В самом общем виде звуки делят на тоны и шумы. Тоны — это гармонические колебания. Они содержат основную частоту и обертоны, отличающиеся от основной в целое число раз. Шумы состоят из частот, не находящихся в гармонических отношениях. Анализ частот звука — одно из важнейших функций слуховой системы.
78


 Психофизические характеристики звука.Слуховой орган (периферический отдел слуховой системы) — это измерительное устройство, которое преобразует физические параметры звука (интенсивность, частоту, длительность) в активность периферических и центральных слуховых нейронов, на основе чего строятся субъективные характеристики звука (громкость, высота, продолжительность). В органе слуха происходит взаимодействие звуковых частот в определенных диапазонах (критические полосы слуха), в слуховых центрах — взаимодействие рецептивных полей, в ощущении — маскировка одного тона на фоне другого (или отсутствие маскировки).
Психофизические характеристики звука.Слуховой орган (периферический отдел слуховой системы) — это измерительное устройство, которое преобразует физические параметры звука (интенсивность, частоту, длительность) в активность периферических и центральных слуховых нейронов, на основе чего строятся субъективные характеристики звука (громкость, высота, продолжительность). В органе слуха происходит взаимодействие звуковых частот в определенных диапазонах (критические полосы слуха), в слуховых центрах — взаимодействие рецептивных полей, в ощущении — маскировка одного тона на фоне другого (или отсутствие маскировки).
Психофизическим эквивалентом интенсивности звука является его громкость. Более интенсивные звуки воспринимаются как более громкие. Однако, между психофизической характеристикой звука — громкостью и его физической интенсивностью нет прямого соответствия. Для измерения громкости применяют метод сравнения стандартного тона определенной частоты с другим тоном той же частоты. Второй тон отличается от первого по интенсивности. Полученные оценки громкости в условиях сравнения двух тонов свидетельствуют о том, что громкость (J) возрастает как кубический корень из интенсивности звука (I) и описывается формулой J=kI0,3. Таким образом, если интенсивность звука увеличивается на 10 дБ, то громкость увеличивается в два раза.
Международная организация по стандартизации ввела единицу громкости — сон, которая представляет собой громкость тона при частоте 1000 Гц и интенсивности 40 дБ над порогом ощущения. Соотношения между интенсивностью звука и ощущением громкости сравниваются на рис. 16.6 с известными из повседневного опыта ощущениями.
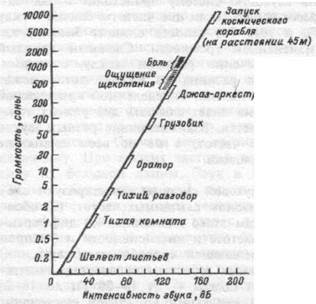
|
| Рис. 16.6. Соотношение интенсивности звука (физическая единица) и его громкости (единица ощущения). |
 79
79
 Громкость звука зависит не только от интенсивности, но и от его частоты. Чувствительность слуха к разным частотам различна. Звуки, интенсивность которых меньше, чем предел чувствительности слуха, не слышны. Порог, то есть едва слышимый звук, наличие которого человек определяет с вероятностью 0,5 можно считать равным по громкости для разных частот. При интенсивностях, выше пороговых, равная громкость достигается при разных для каждой частоты уровнях интенсивности — это кривые равной громкости. Их оценка имеет важное диагностическое значение при нарушениях слуха.
Громкость звука зависит не только от интенсивности, но и от его частоты. Чувствительность слуха к разным частотам различна. Звуки, интенсивность которых меньше, чем предел чувствительности слуха, не слышны. Порог, то есть едва слышимый звук, наличие которого человек определяет с вероятностью 0,5 можно считать равным по громкости для разных частот. При интенсивностях, выше пороговых, равная громкость достигается при разных для каждой частоты уровнях интенсивности — это кривые равной громкости. Их оценка имеет важное диагностическое значение при нарушениях слуха.
Для того, чтобы определить интенсивности разных тонов, вызывающих ощущение равной громкости, пользуются методом сравнения стандартного тона частотой 1000 Гц, интенсивностью 40 дБ и длительность 0,5 с тоном другой частоты той же длительности. Слушатель должен сам подобрать такую интенсивность сравниваемого тона, при которой его громкость будет такая же, как громкость стандартного тона. Чем больше интенсивность тонов, тем меньше их различия по громкости. Последняя "выравнивается" при возрастании интенсивности звука. В психофизических единицах — сонах — измеряют громкость не только чистых тонов, но и сложных звуков.
Психофизическим эквивалентом частоты тона является его высота, единицей которой считается мел. В соответствии с общепринятым определением, тон частотой 1000 Гц при 60 дБ имеет высоту 1000 мелов.
Субъективным эквивалентом формы звуковой волны является тембр звука. Если звуковая волна имеет чисто синусоидальную форму, то звук определяется как слабоокрашенный "чистый тон". Если звук содержит много гармоник основной частоты, он воспринимается как более или менее "окрашенный" тон.
Действие двух тонов на слуховую систему происходит в так называемой критической полосе частот. Если две частоты близки между собой, например, 1000 Гц и 1010 Гц, человек слышит биения, т.е. звук с периодическим изменением громкости. Сложный хриплый звук человек слышит приувеличении разницы между частотами, например, 1000 и 1100 Гц. Если разница по шкале частот между звуками увеличивается, то по достижении определенной критической величины этой разницы человек начинает слышать два разных тона, а общая громкость звука уменьшается. Взаимовлияние разных звуков возможно в критической полосе частот, а не во всем диапазоне частот, воспринимаемых ухом человека.
Показатели слуха.Функции слуховой системы характеризуют следующими показателями: (1) диапазоном слышимых частот; (2) абсолютной чувствительностью к звукам этого диапазона; (3) дифференциальной чувствительностью по частоте и интенсивности и (4) пространственной и временной разрешающей способностью слуха.
Диапазон частот, воспринимаемых взрослым человеком, охватывает около 10 октав музыкальной шкалы — от 16-20 Гц до 16-20 кГц. Этот диапазон, характерный для людей до 25 лет, постепенно
80
 уменьшается из года в год за счет сокращения его высокочастотной части. После 40 лет верхняя частоты слышимых звуков уменьшается на 80 Гц каждые последующие полгода.
уменьшается из года в год за счет сокращения его высокочастотной части. После 40 лет верхняя частоты слышимых звуков уменьшается на 80 Гц каждые последующие полгода.
Наибольшая чувствительность слуха имеет место на частотах от 1 до 4 КГц. В этом диапазоне частот чувствительность слуха человека близка к уровню броуновского шума — 2 • 10-5 Па. Судя по аудиограмме, т.е. функции зависимости порога слухового ощущения от частоты звука, чувствительность к тонам ниже 500 Гц неуклонно снижается: на частоте 200 Гц — на 35 дБ, а на частоте 100 Гц — на 60 дБ. Подобное ухудшение чувствительности слуха, на первый взгляд, кажется странным, поскольку оно затрагивает именно тот диапазон частот, в котором лежит большая часть звуков речи и музыкальных инструментов. Однако, было подсчитано, что в пределах области слухового восприятия человек ощущает около 300 000 различных по силе и высоте звуков. Малая чувствительность слуха к звука низкочастотного диапазона предохраняет человека от постоянно ощущения низкочастотных колебаний и шумов собственного тела (движения мышц, суставов, шум крови в сосудах).
Дифференциальная чувствительность слуха человека характеризует способность отличать минимальные изменения параметров звука (интенсивности, частоты, длительности и т.д.). В области средних уровней интенсивностей (порядка 40- 50 дБ над порогом слышимости) и частот 500-2000 Гц дифференциальный порог по интенсивности составляет всего 0.5-1.0 дБ, по частоте 1%. Различия сигналов по длительности, которые воспринимаются . слуховой системой, составляют величины менее 10%, а изменение угла расположения источника высокочастотного тона оценивается с точностью до 1-3°.
Пространственный слух не только позволяет установить место расположения источника звучащего объекта, степень его удаленности и направление его перемещения, но и увеличивает четкость восприятия. Простое сравнение монофонического и стереофонического прослушивания стереофонической записи дает полную картину преимущества пространственного восприятия.
Временные характеристики пространственного слуха базируются на объединении данных, получаемых от двух ушей (бинауральный слух). Бинауральный слух определяют два основных условия. Для низких частот основным фактором является различие во времени попадания звука в левое и правое ухо, для высоких частот — различия в интенсивности. Сначала звук достигает уха, расположенного ближе к источнику. При низких частотах звуковые волны "огибают" голову в силу их большой длины. Звук в воздушной среде имеет скорость 330 м/с. Следовательно, 1 см он проходит за 30 мкс. Поскольку расстояние между ушами человека составляет 17-18 см, а голову можно рассматривать как шар с радиусом 9 см, то разница между попадание звука в разные уши составляет 9п • 30=840 мкс, где 9п (или 28 см) — это тот дополнительный путь, который должен пройти звук вокруг головы, чтобы попасть в другое ухо. Естественно, эта разница зависит от места расположения источника — если он находится по средней линии впереди (или сзади), то звук достигает
81


 обоих ушей одновременно. Малейший сдвиг вправо или влево от средней линии (даже менее 3°), уже воспринимается человеком. А это значит, что значимая для анализа мозгом разница между приходом звука на правое и левое ухо составляет меньше 30 мкс. Следовательно, физическая пространственная размерность воспринимается за счет уникальных способностей слуховой системы как анализатора времени.
обоих ушей одновременно. Малейший сдвиг вправо или влево от средней линии (даже менее 3°), уже воспринимается человеком. А это значит, что значимая для анализа мозгом разница между приходом звука на правое и левое ухо составляет меньше 30 мкс. Следовательно, физическая пространственная размерность воспринимается за счет уникальных способностей слуховой системы как анализатора времени.
Для того, чтобы можно было отметить такую небольшую разницу во времени, необходимы очень тонкие и точные механизмы сравнения. Такое сравнение осуществляется центральной нервной системой в местах, где импульсация от правого и левого ушей сходится на одной структуре (нервной клетке). Подобных мест, так называемых основных уровней конвергенции, в классической слуховой системе не менее трех — это верхнеоливарный комплекс, нижний холм и слуховая кора. Дополнительные места конвергенции находятся внутри каждого уровня, например, межхолмовые и межполушарные связи.
Фаза звуковой волны связана с различиями во времени поступления звука в правое и левое ухо. Более "поздний" звук отстает по фазе от предыдущего, более "раннего" звука. Это отставание имеет значение при восприятии относительно низких частот звуков. Это частоты с длиной волны не менее 840 мкс, т.е. частоты не более 1300 Гц.
При высоких частотах, когда величина головы значительно больше длины звуковой волны, последняя не может "огибать" это препятствие. Например, если звук имеет частоту 100 Гц, то длина его волны составляет 33 м, при частоте звука 1000 Гц — 33 см, а при частоте 10000 Гц — 3,3 см. Из приведенных цифр следует, что при высоких частотах звук отражается головой. В результате' возникает разница и в интенсивности звуков, поступающих на правое и левое ухо. У человека дифференциальный порог по интенсивности на частоте 1000 Гц составляет величину порядка 1 дБ, поэтому оценка местоположения источника звука высокой частоты основывается на различиях интенсивности звука, попадающего на правое и левое ухо.
Разрешающая способность слуха по времени характеризуется двумя показателями. Во-первых, это временная суммация. Характеристики временной суммации — 1) время, в течение которого длительность стимула влияет на порог ощущения звука, 2) степень этого влияния, т.е. величина изменения порога реакции. У человека временная суммация длится около 150 мс. Во-вторых, это минимальный интервал между двумя короткими раздражителями (звуковыми импульсами), которой различается ухом. Его величина составляет 2-5 мс.
Периферический отдел слуховой системы.Периферический отдел — орган слуха, состоит из наружного, среднего и внутреннего уха (рис. 16.7). Несмотря на значительные размеры, наружные структуры уха человека играют небольшую роль в процессах восприятия звука. Функции наружного уха (ушная раковина, наружный слуховой
82

 проход и внешняя сторона барабанной перепонки) сводятся к обеспечению направленного приема звуковых волн. Ушные раковины являются рупором и способствуют концентрации звуков, исходящих из разных участков пространства в направлении наружного слухового прохода, а также ограничивают поток звуковых сигналов, поступающих с тыльной стороны головы.
проход и внешняя сторона барабанной перепонки) сводятся к обеспечению направленного приема звуковых волн. Ушные раковины являются рупором и способствуют концентрации звуков, исходящих из разных участков пространства в направлении наружного слухового прохода, а также ограничивают поток звуковых сигналов, поступающих с тыльной стороны головы.
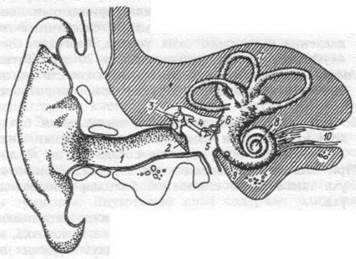
Рис. 16.7. Схема наружного, среднего и внутреннего уха.
1 — слуховой проход, 2 — барабанная препонка, 3 — молоточек, 4 — наковальня, 5 — стремечко, 6 — овальное окно, 7 — полукружные каналы, 8 — улитка, 9 — круглое окно, 10 — слуховой нерв.
Собственная (резонансная) частота колебаний наружного слухового прохода составляет около 3 кГц. Это значит, что коэффициент усиления интенсивности звука имеет максимальную величину именно для тонов этой частоты. Усиление интенсивности звуков на резонансной частоте наружного уха человека заметно на частотно-пороговой кривой слуха и составляет около 10 дБ. Таким образом, минимальный порог слышимости тонов связан с физиологической ролью акустических свойств наружного уха.
Структуры наружного уха несут также защитную функцию. Они охраняют барабанную перепонку от механических и термических воздействий, обеспечивают постоянную температуру и влажность в этой области. Наружный слуховой проход заканчивается барабанной перепонкой, которая передает колебания воздуха в наружном ухе системе косточек среднего уха.
Барабанная перепонка, плошадь которой составляет 66-69,5 мм2, является границей между наружным и средним ухом и имеет форму конуса с вершиной, направленной в полость среднего уха. Ее ос-
83

 новная задача — передача звуковых колебаний в среднее ухо. Среднее ухо соединяется с задней частью глотки узким каналом — евстахиевой трубой. Функция евстахиевой трубы — уравнивание давления в среднем ухе с давлением наружной воздушной среды.
новная задача — передача звуковых колебаний в среднее ухо. Среднее ухо соединяется с задней частью глотки узким каналом — евстахиевой трубой. Функция евстахиевой трубы — уравнивание давления в среднем ухе с давлением наружной воздушной среды.
Колебания барабанной перепонки приводят в движение молоточек (ручка которого прикреплена к барабанной перепонке), присоединяющуюся к молоточку наковальню и конечную в этой цепи косточку — стремечко (рис. 16.7). Основание стремечка, укрепленное в овальном окне улитки, в свою очередь, приводит в движение пе-рилимфу, заполняющую вестибулярный и барабанный ход улитки. Звуковое давление у круглого окна улитки, благодаря передаточной функции слуховых косточек, усиливается в 20 раз. Такое усиление несет большую функциональную роль, т.к. жидкость внутреннего уха обладает значительно большим акустическим сопротивлением, чем воздух.
Среднее ухо человека имеет полосу пропускания без ослабления сигналов до 1 кГц. Наклон частотной характеристики фильтра среднего уха на более высоких частотах составляет от 7 до 12 дБ на октаву. При высоких интенсивностях звука коэффициент передачи среднего уха также резко снижается, благодаря сокращению мышц среднего уха.
В среднем ухе имеются две мышцы: мускул, натягивающий барабанную перепонку и прикрепленный к ручке молоточка, и стапеди-альный мускул, прикрепленный к стремечку. Функция этих мышц состоит в том, что сокращаясь, они уменьшают амплитуду колебаний барабанной перепонки и косточек и, тем самым, снижают коэффициент передачи уровня звукового давления во внутреннее ухо. Сокращения мышц возникают при больших интенсивностях звуков — больше 90 дБ, и несут защитную функцию. Латентный период сокращения слишком велик (порядка 10 мс), чтобы предохранить ухо от действия резких внезапных звуков. Но при длительном пребывании в условиях действия сильных шумов сокращение мышц приобретает важную защитную роль. Сокращения мышц, особенно стапедиального мускула, происходят также при действии нового акустического раздражителя, при глотании, жевании и зевании, а также при речевой деятельности. Это свидетельствует о том, что мышцы среднего уха участвуют не только в защитном акустическом рефлексе, но также в ориентировочной реакции и в реализации обратной связи от речевой системы к слуховому входу: когда человек говорит или поет, стапедиальный мускул сокращается и низкочастотные звуки подавляются, а высокочастотные проходят среднее ухо без искажений.
Важнейшую функцию рецепции звука несет улитка — костная структура внутреннего уха, закрученная в виде спирали. При попадании в ухо звуковой волны, приводящей в движение барабанную перепонку, а затем цепь слуховых косточек среднего уха, основание стремечка вдавливает эластичную мембрану овального окна, передавая давление в полость улитки через движение жидкости — пери-лимфы.
84


 Внутри улитки, по всей ее длине, проходят две мембраны — основная и рейснерова, разделяющие улитку на три части, так называемые лестницы. Это вестибулярная, барабанная и средняя лестницы, заполненные несжимаемыми жидкостями. Средняя лестница, в которой находится рецепторный аппарат — орган Корти, заполнена эндолимфой. Эндолимфа средней лестницы сообщается с эндолимфой вестибулярного органа и имеет тот же состав. Вестибулярная и барабанная лестницы заполнены перилимфой — внеклеточной жидкостью иного состава, чем эндолимфа. Состав эндолим-фы обеспечивает секреторная функция сосудистой полоски, расположенной на наружной стенке средней лестницы.
Внутри улитки, по всей ее длине, проходят две мембраны — основная и рейснерова, разделяющие улитку на три части, так называемые лестницы. Это вестибулярная, барабанная и средняя лестницы, заполненные несжимаемыми жидкостями. Средняя лестница, в которой находится рецепторный аппарат — орган Корти, заполнена эндолимфой. Эндолимфа средней лестницы сообщается с эндолимфой вестибулярного органа и имеет тот же состав. Вестибулярная и барабанная лестницы заполнены перилимфой — внеклеточной жидкостью иного состава, чем эндолимфа. Состав эндолим-фы обеспечивает секреторная функция сосудистой полоски, расположенной на наружной стенке средней лестницы.
Основная мембрана имеет в развернутом виде около 3,5 см в длину, а ширина ее возрастает по направлению от овального окна к вершине. На основной мембране находится скопление чувствительных рецепторных клеток, входящих в состав органа Корти фо-норецепторов. Это — механореиепторы, представленные у позвоночных животных и человека волосковыми клетками. Количество этих клеток, каждая из которых имеет до сотни волосков, составляет у человека около 25 тысяч. Волосковые клетки располагаются в два слоя, разделенные между собой каналом небольшого размера — кортиевым туннелем. Внутренний слой содержит один ряд, а наружный — 3-5 рядов клеток. Обшее число наружных клеток достигает почти 20 тысяч, а внутренних — около 3,5-5 тысяч. Волосковые клетки прикрыты сверху покровной, или текториальной, мембраной соединительнотканного происхождения. У покровной мембраны закреплен только один край, второй свободен. Поэтому она скользит по структурам, расположенным под ней,- особенно когда двигается основная мембрана при передаче звука во внутреннем ухе. Покровная мембрана сгибает волоски рецепторных клеток, погруженные в ее вещество.
Движение основной мембраны с расположенными на ней рецептор-ным аппаратом вызывает деформацию волосков волосковых клеток органа Корти, причем воздействие на наружные волосковые клетки оказывается сильнее, чем на внутренние, поскольку основная мембрана закреплена. В результате деформации волосков возникает активность рецепторных клеток и связанных с ними нервных окончаний.
Волокна основной мембраны улитки внутреннего уха настроены на колебания различных звуковых частот: лежащие у основания улитки резонируют при воздействии высоких частот, а лежащие у ее вершины — низких частот. Место максимального отклонения мембраны связано с частотой звука: для звуков высокой частоты оно расположено у овального окна, для звуков низкой частоты — у вершины улитки. В результате, разные звуковые частоты преобразуются в амплитуды отклонения мембраны, локализованные в разных ее частях. Однако, место максимального отклонения мембраны улитки, приводящее к возбуждению отдельных локально расположенных групп рецепторных волосковых клеток, недостаточно для восприятия информации о звуке. На уровне рецепторных клеток происходит трансформация механического процесса в электрический,
85


 преобразование акустических сигналов, поступающих из внешней среды, в формы активности, присущие нервной системе — медленные электрические потенциалы и короткие импульсы.
преобразование акустических сигналов, поступающих из внешней среды, в формы активности, присущие нервной системе — медленные электрические потенциалы и короткие импульсы.
В улитке существует три типа электрических потенциалов: 1) микрофонный; 2) суммационный; 3) эндокохлеарный.
Микрофонный потенциал представляет собой результат деформации волосковых клеток Кортиева органа. Этот потенциал следует частоте звуковых стимулов до 4000- 5000 Гц, возрастает по величине с увеличением интенсивности звука. Свое название микрофонный потенциал получил благодаря тому, что он, подобно микрофону, полностью воспроизводит форму и частоту звуковых волн. При частотах стимула, превышающих 4000- 5000 Гц, проявляется суммационный потенциал — сдвиг постоянного потенциала улитки, слабо выраженный при низких частотах. Величина этого потенциала, так же как микрофонного, пропорциональна интенсивности звука. Микрофонный и суммационный потенциал являются рецепторными потенциалами органа слуха, при этом микрофонные потенциалы считаются результатом электрического возбуждения рецепторных клеток, а суммационные — окончаний волокон слухового нерва — дендритов биполярных клеток спирального ганглия, распределенных между волосковыми клетками улитки. Эндокохлеарный (или эндолимфати-ческий) потенциал — постоянный потенциал улитки. Его источник — активный секреторный процесс сосудистой полоски, которая выделяет эндолимфу. Величина эндокохлеарного потенциала в эндо-лимфе на 80 мВ больше, чем средняя величина потенциала в любой части организма. Верхушка рецепторных клеток, покрытая волосками, омывается эндолимфой, богатой калием и бедной натрием. Их мембрана поддерживает крутой градиент потенциала, т.к. она отделяет эндолимфу с зарядо
Дата добавления: 2016-03-27; просмотров: 2206;