Регуляция слюноотделения
Прием пищи возбуждает слюноотделение рефлекторно. При достаточно сильном раздражении и высокой возбудимости пищевого центра слюноотделение начинается через 1—3 с, а при слабой силе раздражения латентный период достигает 20—30 с. Слюноотделение продолжается весь период еды и прекращается вскоре после нее.
От рецепторов полости рта сигналы передаются в ЦНС по афферентным волокнам тройничного, лицевого, языкоглоточного и блуждающего нервов. Основной слюноотделительный центр расположен в продолговатом мозге. Именно сюда, а также в боковые рога верхних грудных сегментов спинного мозга поступают сигналы из полости рта и расположенных выше отделов мозга. Отсюда влияния по эфферентным парасимпатическим и симпатическим нервным волокнам направляются к слюнным железам.
Парасимпатическая иннервация слюнных желез начинается из ядер продолговатого мозга (рис. 179). Симпатическая иннервация слюнных желез осуществляется от боковых рогов II—IV грудных сегментов спинного мозга.
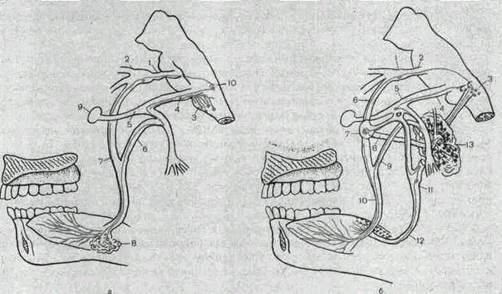
Рис.179. Схема путей рефлекторного возбуждения подчелюстной (а) и околоушной (б) слюнных желез.
а: 1 —тройничный нерв; 2—узел тройничного нерва (Гассеров); 3—ядро лицевого нерва; 4 — лицевой нерв; 5 — коленчатый узел; 6 — барабанная струна; 7 — язычный нерв; 8 — подчелюстная железа и подчелюстной ганглий; 9 — крылонебный ганглий; 10 — центр слюноотделения, б: 1 — тройничный узел; 2 — тройничный нерв; 3 — ядро языкоглоточного нерва; 4 -^ лицевой нерв; 5 — коленчатый узел; 6 — верхнечелюстной нерв; 7 — ушной узел; 8 — ушно-височный нерв; 9 — барабанная струна; 10 — язычный нерв; 11 — пещеристый узел; 12—языкоглоточный нерв; 13—околоушная слюнная железа. Выделены чувствительные нервные волокна, идущие от рецепторных аппаратов языка, и преганглионарные парасимпатические волокна;
пунктирная линия— постганглионарные парасимпатические волокна.
В окончаниях постганглионарных парасимпатических волокон высвобождается медиатор — ацетилхолин, возбуждающий секреторные клетки слюнных желез. При этом выделяется большое количество жидкой слюны. Длительное раздражение парасимпатических нервов вызывает истощение слюнных желез: снижается содержание в слюне органических компонентов, в гландулоцитах резко уменьшается количество гранул.
В окончаниях симпатических нервных волокон высвобождается норадреналин. Под его влиянием выделяется небольшое количество густой слюны. Раздражение симпатических нервов усиливает образование гранул секрета в гландулоцитах. Одновременное или предшествующее раздражение парасимпатических нервов усиливает симпатические секреторные эффекты. Ответная реакция при раздражении симпатических нервов возникает спустя более длительный латентный период, чем после раздражения парасимпатических.
Различия в секреторной деятельности слюнных желез в ответ на прием различных видов пищи можно объяснить модуляцией частоты импульсов по парасимпатическим и симпатическим нервным волокнам и изменением соотношении между их влияниями. Усиление слюноотделения сопровождается расширением кровеносных сосудов железы, что повышает уровень ее кровоснабжения. Сосудорасширяющие эффекты в железе вызываются парасимпатическими нервными волокнами, образующимися в ней продуктами обмена веществ, а также кининами. Полагают также, что определенную роль играет уменьшение сопротивления сосудов в результате освобождения железы от некоторого количества секрета.
Слюноотделение начинается по типу условных рефлексов — в ответ на вид и запах пищи.
Рефлекторные влияния могут и тормозить слюноотделение, вплоть до его прекращения. Такое торможение может быть вызвано болевым раздражением, отрицательными эмоциями, умственным напряжением, дегидратацией организма. Все эти воздействия снижают активность пищевого центра и его части — центра слюноотделения. Возбудителями последнего могут быть некоторые гуморальные вещества. Так, обильное отделение слюны наблюдается при асфиксии вследствие раздражения центра слюноотделения угольной кислотой. Раздражают его некоторые токсины и при этом наблюдается обильное слюноотделение.
Перерезка парасимпатических нервов, иннервирующих слюнные железы, временно резко усиливает выделение ими слюны — паралитическая секреция. В первые три дня непрерывное слюноотделение обусловлено повышенным выделением ацетилхолина вследствие дегенерации нейронов, поэтому эта секреция называется дегенеративной. В последующие дни паралитическая секреция связана с повышенной чувствительностью денервированной железы к ряду веществ, циркулирующих в крови, к которым железа с нормальной иннервацией малочувствительна. Холиномиметические фармакологические вещества вызывают усиление слюноотделения, холинолитические -- тормозят его.
Глотание
Глотание является рефлекторным актом, центр его находится в продолговатом мозге, на дне IV желудочка. /Доказательством рефлекторной природы глотания служит следующее: если смазать корень языка и глотки раствором кокаина и выключить таким образом рецепторы их слизистой оболочки, то глотание осуществляться не будет. Глотание невозможно и после перерезки эфферентных нервов глотки.
Рефлекс глотания состоит из ряда последовательных звеньев. Строгая координация звеньев глотательного рефлекса обеспечивается сложными взаимосвязями различных отделов ЦНС — от продолговатого мозга до коры большого мозга. Центр глотания связан с центром дыхания, что обеспечивает задержку дыхания во время глотания и имеет значение в предотвращении попадания пищи в воздухоносные пути. Связь центра глотания с центром регуляции сердечной деятельности подтверждается учащением сердечных сокращений во время глотания.
Акт глотания делится на три фазы: 1) ротовую (произвольную), 2) глоточную (быструю непроизвольную), 3) пищеводную (медленную непроизвольную). Акт глотания изучен рентгеновским методом (рис. 180).
Из пищевой пережеванной массы во рту отделяется пищевой комок объемом 5—15 см3, который движениями языка и щек перемещается на спинку языка. Сокращениями передней части языка пищевой комок прижимается к твердому небу, затем последовательными сокращениями средней части отжимается кзади и переводится на корень языка за передние дужки (!-я 4'аза).
Раздражение рецепторов слизистой оболочки корня языка рефлекторно вызывает сокращение мышц, приподнимающих мягкое небо, и мышц языка, что препятствует попаданию пищи в полость носа. Движения языка способствуют проталкиванию пищи в полость глотки. Одновременно с этим происходит сокращение мышц, смещающих подъязычную кость и вызывающих поднятие гортани, вследствие чего закрывается вход в дыхательные пути, чти препятствует поступлению в них пищи. Переводу пищи в глотку способствует повышение давления в полости рта. Возвращению пищи из глотки в полость рта препятствуют поднявшийся вверх корень языка и плотно прилегающие к нему дужки. Как только пища поступила в полость глотки, начинают сокращаться мышцы, суживающие просвет глотки выше пищевого комка, вследствие чего последний передвигается в пищевод. Существенное значение при этом имеет градиент давления между полостью глотки и началом пищевода. Перед глотанием глоточно-пищеводный сфинктер закрыт, во время глотания давление в глотке повышается до 6 Па (45 мм рт. ст.) и через открывшийся сфинктер пищевой комок поступает в пищевод (2-я фаза), где давление в это время не более 4 Па (30 мм рт. ст.). Давление в глотке значительно выше, сфинктер закрыт и невозможно обратное забрасывание пищевого комка в глотку. Две фазы глотательного цикла длятся около 1 . 
Рис.180. Последовательность движений в полости рта и глотке при глотании.
Вторую фазу глотания нельзя выполнить произвольно, если в полости рта нет пищи или слюны. В этом легко убедиться, делая подряд несколько глотательных движений: первое движение происходит легко, так как в полости рта всегда имеется небольшое количество слюны; при отсутствии слюны, которая проглочена, дальнейшие глотательные движения осуществить не удается.
Третью фазу глотания составляют прохождение.пищи по пищеводу и перевод ее в желудок. Движения пищевода взаимосвязаны с движениями глотательного аппарата и вызываются рефлек-торно при каждом глотательном акте. Если в эксперименте на собаках перерезать пищевод и вкладывать пищу непосредственно в него, то она передвигается в сторону желудка только после совершения глотательных движений. Продолжительность продвижения по пищеводу твердой пищи в среднем 8—9 с, жидкой 1—2 с.
В момент акта глотания пищевод подтягивается к зеву и начальная часть его расширяется, принимая пищевой комок.
Сокращения пищевода имеют характер волны, возникающей в верхней его части и распространяющейся в сторону желудка. При этом последовательно сокращаются кольцеобразно расположенные мышцы пищевода (в верхней трети поперечнополосатые, в нижних двух третях — гладкие), передвигая перед волной сокращения находящийся в несколько расширенной части пищевода пищевой комок в сторону желудка. Такой тип сокращений называется перистальтическим.
Первичная перистальтическая волна, вызываемая актом глотания, доходит до уровня пересечения пищевода с дугой аорты. Далее возникает вторичная перистальтическая волна, вызываемая не актом глотания, а первичной перистальтической волной. Вторичная волна продвигает пищевой комок до кардиальной части желудка. Средняя скорость распространения перистальтической волны по пищеводу человека 2—5 см/с, она в большой мере зависит от свойств пищи. Вторичная перистальтическая волна может быть вызвана остатком пищевого комка в нижней трети пищевода, благодаря чему этот остаток переводится в желудок.
Регуляция моторной деятельности пищевода осуществляется в основном эфферентными волокнами блуждающего и симпатического нервов; определенная роль принадлежит интрамуральным нервным образованиям пищевода.
Вне глотательных движений вход из пищевода в желудок закрыт. Когда пища и перистальтическая волна достигают конечной части пищевода, происходит рефлекторное понижение тонуса мышц кардии и пищевой комок переходит в желудок. При наполнении желудка тонус кардии повышается, что предупреждает забрасывание содержимого желудка в пищевод. Парасимпатические волокна блуждающего нерва стимулируют перистальтику пищевода и расслабляют кардию, симпатические волокна тормозят моторику пищевода, но повышают тонус кардиальной частив
ПИЩЕВАРЕНИЕ В ЖЕЛУДКЕ
Желудок принимает участие в пищеварительном конвейере, а также в межуточном обмене веществ, поддержании постоянства рН крови и кроветворении (вырабатывает желудочный мукопротеид, или внутренний фактор Касла). Пищеварительными функциями желудка являются: депонирование пищи, ее механическая и химическая обработка, постепенная порционная эвакуация пищевого содержимого в кишечник. Находясь в желудке в течение нескольких часов, пища набухает, разжижается, многие ее компоненты растворяются и подвергаются гидролизу ферментами слюны и желудочного сока. Желудочный сок обладает также антибактериальным действием.
Карбогидразы слюны продолжают действовать на углеводы пищи, находящиеся в центральной части пищевого содержимого желудка (куда еще не диффундировал кислый желудочный сок, прекращающий действие карбогидраз слюны). Ферменты кислого желудочного сока воздействуют на пищевые белки в относительно узкой зоне пищевого содержимого, находящегося в непосредственном контакте со слизистой оболочкой желудка и в небольшом удалении от нее, куда диффундировал желудочный сок и не был нейтрализован за счет буферных свойств пищи. Ширина этой зоны зависит от количества и свойств желудочного сока и принятой пищи. Вся масса пищи в желудке не смешивается с соком. По мере разжижения и химической обработки пищи ее слой, прилегающий к слизистой оболочке, движениями желудка перемещается в его антральную часть, откуда эвакуируется в кишечник. Таким образом, пищеварение в желудке (по типу полостного) осуществляется некоторое время за счет слюны, но ведущее значение имеют секреторная и моторная деятельность самого желудка.
Дата добавления: 2016-03-27; просмотров: 1897;