Рефлекторная деятельность заднего мозга
Задний мозг выполняет сложные и многообразные функции, многие из которых являются жизненно важными для организма.
Рефлекторные соматические реакции направлены на поддержание позы, на обеспечение восприятия, обработки и проглатывания пищи.
В обеспечении последних процессов принимают участие также и вегетативные рефлексы, регулирующие секрецию слюнных желез и других пищеварительных желез.
Рефлексы, направленные на поддержание позы, в первую очередь связаны с рецепторами вестибулярного аппарата и полукружных каналов. Различают две группы рефлексов позы: статические и статокинетические.
Статические рефлексы в свою очередь разделяются на рефлексы положения и рефлексы выпрямления. Рефлексы положения обеспечивают изменение тонуса мышц при перемене положения тела в пространстве. Рефлексы выпрямления определяют перераспределение тонуса мышц, приводящее к восстановлению естественной для данного вида животного позы в случае ее изменения. В осуществлении этих рефлексов принимают участие вестибулярные афферентные волокна и нейроны латерального вестибулярного ядра, аксоны которых идут в спинной мозг в составе вестибулоспинального тракта. Рефлекторная дуга указанных рефлексов включает в себя небольшое число последовательно включенных нервных элементов, что обеспечивает эффективную и своевременную коррекцию позы при вестибулярных раздражениях благодаря моносинаптическим связям быстропроводящих вестибулоспинальных волокон с мотонейронами мышц-разгибателей и параллельному торможению мотонейронов мышц-сгибателей.
Более сложный характер имеют вестибулярные рефлексы выпрямления, основной компонент которых представлен рефлекторными влияниями на мышцы шеи. Благодаря перераспределению тонуса шейных мышц голова постоянно сохраняет нормальное положение.
Наиболее сложный характер имеют статокинетические рефлексы, направленные на сохранение позы и ориентацию в пространстве при изменении скорости движения.
Так, при внезапной остановке вагона трамвая или автобуса, или при крутом повороте автомобиля, происходят сокращения мышц, направленные на преодоление действующих на человека ускорений. Развивающиеся двигательные реакции отличаются значительной силой, быстротой и сложностью, представляя собой резкие фазные ответы, отличные от медленных позных реакций, характерных для статических рефлексов.
В наш механизированный век с его необычно высокими скоростями статокинетические рефлексы могут играть не только положительную (защитную), но и отрицательную роль. Например, водитель автомобиля может потерять координацию в момент неожиданного удара не замеченным на дороге предметом. Быстрое рефлекторное разгибание рук и ног приведет, с одной стороны, к неверному повороту руля, а с другой — к ненужному нажиму на педаль акселератора. В результате этого автомобиль может получить новые неожиданные толчки в такой последовательности, что водитель не успеет возвратиться в состояние равновесия. В этом случае правильнее будет сказать, что не автомобиль, а мышцы водителя вышли из-под контроля в результате срабатывания статокинетических рефлексов. Эти рефлексы, так же как и статические, вовлекают в деятельность почти всю мускулатуру тела. Особенно сильно они выражены на глазных мышцах. Движение глазных мышц обеспечивает сохранение нормальной зрительной ориентации при ускорении или замедлении движения и носит название «нистагм».
Кроме двигательных рефлексов, активация вестибулярного аппарата возбуждает и вегетативные центры, в том числе ядра блуждающего нерва. Возникающие при этом вестибуловегетативные рефлексы приводят к изменениям со стороны дыхания, частоты сердечных сокращений, деятельности желудочно-кишечного тракта.
Большая группа двигательных рефлексов, протекающих при участии ядер заднего мозга, связана с процессами, обеспечивающими захват, пережевывание и проглатывание пищи. Характерным для этих рефлексов является их связь между собой, благодаря чему осуществление одного рефлекса вызывает включение последующего («цепные рефлексы») . В пищевых двигательных рефлексах принимают участие двигательные ядра тройничного, языкоглоточного, блуждающего, добавочного и подъязычного нервов. Мотонейроны тройничного нерва обеспечивают рефлекс жевания. Глотание пищи и продвижение ее в пищевод обеспечивается цепным рефлексом, последовательно вовлекающим нейроны ядер языкоглоточного, блуждающего, добавочного и подъязычного нервов. Рецептивное поле этих рефлексов составляют рецепторы слизистой оболочки рта, корня языка.
Вегетативные ядра заднего мозга относятся к парасимпатическому отделу нервной системы и обеспечивают различные вегетативные рефлексы. В пределах продолговатого мозга и моста мозга локализуются преганглионарные эфферентные нейроны, аксоны которых поступают в периферические вегетативные ганглии.
Основные вегетативные ядра продолговатого мозга входят в систему блуждающего нерва. Благодаря активации нейронов этих ядер, а также нейронов вегетативных ядер лицевого и языкоглоточного нервов осуществляется рефлекторный контроль дыхания, деятельности сердца, тонуса сосудов, функции пищеварительных желез. Важной особенностью центра, регулирующего дыхание, является то, что он активируется не только импульсами, приходящими по афферентным путям, но и химическими раздражителями.
Нервные клетки дыхательного центра находятся в ретикулярной формации в области IV желудочка. Повреждение этой зоны приводит к остановке дыхания. Исследования Н. А. Миславского, проведенные еще в конце прошлого века, показали, что в дыхательном центре можно выделить две зоны. Одна из них связана преимущественно с осуществлением акта вдоха, другая — с актом выдоха. Поэтому в дыхательном центре можно различать инспираторный и экспираторный полуцентры, которые в общей сложности захватывают ретикулярную формацию до верхней границы моста мозга. Подробное описание структуры и функции дыхательного центра дано в главе 11.
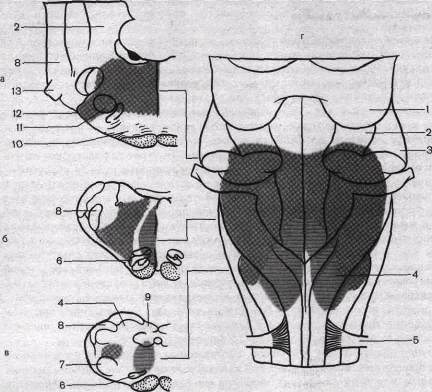
Рис. 86. Локализация различных зон сосудодвигательного центра в продолговатом мозге кошки.
Прессорные зоны выделены перекрестной штриховкой, депрессорные — горизонтальной; а, б, в — поперечные срезы продолговатого мозга; г—дорсальная поверхность. 1—нижние бугры четверохолмия; 2—ножки моста; 3—brachium conjunctivum; 4—нежное ядро; 5—первый шейный корешок; 6—нижнее оливарное ядро; 7—латеральное ретикулярное ядро; 8—спинальный тригеминальный тракт; 9—клиновидное ядро;
10 — трапециевидное тело; 11 — верхнее оливарное ядро; 12 — лицевой нерв; 13 — тройничный нерв.
Второй жизненно важной системой ретикулярной формации продолговатого мозга и моста являются центры, регулирующие деятельность сердца и тонус сосудов. Раздражение одних участков ретикулярной формации дна IV желудочка вызывает увеличение тонуса сосудов и повышение артериального давления, раздражение других — расширение сосудов и падение артериального давления (рис. 86). Таким образом, существуют нейроны, активация которых вызывает сосудорасширяющий или сосудосужающий эффект. Этим клеткам также свойственна постоянная тоническая деятельность, которая может модулироваться изменением афферентной импульсации, идущей от сердца и сосудов. Кроме того, для нейронов сосудодвигательного центра свойственна высокая химическая чувствительность. Вследствие этого частота их ритма определяется изменениями химического состава крови.
Таким образом, центры продолговатого мозга регулируют деятельность многих органов грудной и брюшной полости и другие вегетативные функции. Наличие здесь дыхательного и сердечно-сосудистого центров объясняет тот факт, что нормальное функционирование этого отдела U.HC является жизненно необходимым. В то время, как повреждение других отделов ЦНС может протекать бессимптомно (вследствие больших компенсаторных возможностей мозга), малейшие повреждения (травма, сдавливание опухолью, отек мозга, кровоизлияния) этой области, как правило, немедленно приводят к тяжелым нарушениям жизнедеятельности и даже к смерти.
Дата добавления: 2016-03-27; просмотров: 2724;