РЕФЛЕКТОРНАЯ ДЕЯТЕЛЬНОСТЬ СПИННОГО МОЗГА
В спинном мозге замыкается огромное количество рефлекторных дуг, с помощью которых регулируются как соматические, так и вегетативные функции организма. К ч.ислу наиболее простых рефлекторных реакций относятся сухожильные рефлексы и рефлексы растяжения, вызываемые раздражением рецепторов растяжения той же мышцы, которая развивает рефлекторное сокращение. Центральные окончания афферентных волокон от рецепторов растяжения образуют синапсы непосредственно на мотонейронах без дополнительных переключении на вставочных нейронах. Таким образом, дуга этих рефлексов может иметь моносинаптический характер. Указанное обстоятельство, а также высокая скорость проведения по афферентным волокнам, идущим от мышечных рецепторов и по аксонам ос-мотонейронов, обеспечивают короткое время рефлекса (что особенно демонстративно в случае сухожильных рефлексов).
Сухожильные рефлексы легко вызываются с помощью короткого удара по сухожилию и имеют важное диагностическое значение в неврологической практике. Рефлекторная реакция проявляется в виде резкого сокращения мышцы. Особенно выражены сухожильные рефлексы в мышцах разгибателей ноги, таких, как четырехглавая мышца бедра (коленный рефлекс) или трехглавая мышца голени (ахиллов рефлекс). Однако сухожильные рефлексы вызываются и в мышцах-сгибателях. На руке они четко проявляются на двуглавой и трехглавой мышцах, на лице — на мышцах нижней челюсти.
Быстрота мышечного сокращения и отсутствие последействия обусловлены способом вызывания сухожильного рефлекса. Адекватным раздражителем для соответствующих рецепторов является растяжение мышцы. Постукивание по сухожилию растягивает мышцу только на очень краткий срок.
Зато при этом чувствительные к растяжению рецепторы активируются с высокой степенью синхронности.
Поскольку афферентные волокна, идущие в спинной мозг от рецепторов растяжения, представляют собой довольно гомогенную группу по диаметру и скоростям проведения, афферентные импульсы поступают к мотонейронам в виде синхронной волны. В результате мотонейроны отвечают с незначительной временной дисперсией, посылая в двигательный нерв синхронный разряд, вызывающий короткое мышечное подергивание, сходное с ответом мышцы на одиночное электрическое раздражение двигательного нерва.
Совсем иначе характеризуется рефлекс растяжения, возникающий при адекватном раздражении тех же самых мышечных рецепторов. Естественные растяжения обычно прикладываются к мышцам под действием силы тяжести. Так, при стоянии четырехглавая мышца бедра подвергается растяжению из-за тенденции колена сгибаться под влиянием гравитационных сил. Возникающая в ответ на это растяжение афферентная импульсация характеризуется значительной асинхронностью, так как многочисленные рецепторы растяжения под влиянием постоянной нагрузки генерируют ритмические импульсы, частота которых определяется индивидуальным порогом каждого рецептора. Мотонейроны получают длительные асинхронные импульсы и сами разряжаются асинхронно. В результате этого мышца отвечает плавным длительным сокращением, автоматически противодействующим силе тяжести. Это определяет большое физиологическое значение рефлекса растяжения как механизма поддержания выпрямленной позы или стояния.
Более сложно организованы рефлекторные ответы, выражающиеся в координированном сгибании или разгибании мышц конечности. Сгибательные рефлексы направлены на избежание различных повреждающих воздействий. Поэтому рецептивное поле сгибательного рефлекса достаточно сложно и включает различные рецепторные образования и различные по скорости проведения афферентные пути.
Сгибательный рефлекс возникает при раздражении болевых рецепторов кожи, мыщц и внутренних органов. Вовлекаемые при этих раздражениях афферентные волокна имеют широкий спектр скоростей проведения — от миелинизированных волокон группы А до немиелинизированных волокон группы С. Все разнообразные афферентные волокна, импульсация по которым приводит к развитию сгибательного рефлекса, объединяют под названием афферентов сгибательного рефлекса.
Центральное время сгибательного рефлекса довольно продолжительно, что обусловлено его полисинаптическим характером. Сгибательные рефлексы отличаются от собственных рефлексов мышц — миостатических и сухожильных не только большим числом синаптических переключении на пути к мотонейронам, но и вовлечением ряда мышц, координированное сокращение которых обусловливает движение целой конечности. Одновременно с возбуждением мотонейронов, иннервирующих мышцы-сгибатели, происходит реципрокное торможение мотонейронов мышц-разгибателей.
|
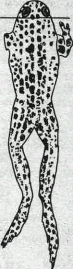
Рис. 83. Сгибательный тонический рефлекс лягушки по Бронджесту.
После декапитации лягушка подвешена за челюсть. На правой стороне перерезаны дорсальные корешки, вследствие чего на этой конечности отсутствует сгибательный тонус.
При достаточно интенсивном раздражении рецепторов задней конечности происходят иррадиация возбуждения и вовлечение в реакцию мышц передней конечности и туловища. При активации мотонейронов противоположной стороны тела наблюдается не сгибание, а разгибание мышц задней конечности — перекрестный разгибательный рефлекс.
К разгибательным рефлексам, кроме уже рассмотренных выше миостатических рефлексов и перекрестного разгибательного рефлекса, относится ряд полисинаптических рефлексов, возникающих при раздражении рецепторов стопы.
Еще более сложный характер имеют ритмические и позные рефлексы, или рефлексы положения. К ритмическим рефлексам у млекопитающих относится чесательный рефлекс. Его аналогом у земноводных является потирательный рефлекс. Ритмические рефлексы характеризуются координированной работой мышц конечностей и туловища, правильным чередованием сгибания и разгибания конечностей наряду с тоническим сокращением приводящих мышц, устанавливающих конечность в определенное положение к кожной поверхности.
Позные рефлексы представляют собой большую группу рефлексов, направленных на поддержание определенной позы, что возможно при наличии определенного мышечного тонуса. Примером позного рефлекса является сгибательный тонический рефлекс, который у лягушки определяет основную позу — сидение. Даже у декапитированной лягушки, подвешанной к штативу, задние конечности всегда несколько согнуты и сгибательный тонус исчезает только после разрушения спинного мозга или перерезки дорсальных корешков. Различия в тонусе сгибательных мышц особенно отчетливы, если перерезку дорсальных корешков осуществить на одной стороне, оставив другую интактной (рис. 83).
Сгибательный тонический рефлекс наблюдается и у млекопитающих, для которых характерно подогнутое положение конечностей (кролик). В то же время для большинства млекопитающих главное значение для поддержания положения тела играет не сгибательный, а разгибательный рефлекторный тонус.
Ввиду того что особенно важную роль в рефлекторной регуляции разгибательного тонуса играют шейные сегменты спинного мозга, специально выделяют шейные тонические рефлексы положения. Эти рефлексы были впервые описаны голландским физиологом Р. Магнусом (1924).
Рецептивным полем шейных тонических рефлексов являются проприорецепторы мышц шеи и фасций, покрывающих шейный участок позвоночника. Центральная часть рефлекторной дуги имеет полисинаптический характер, т. е. включает вставочные нейроны.
Рефлекторная реакция вовлекает мышцы туловища и конечностей. Кроме спинного мозга, в ней участвуют и моторные ядра мозгового ствола, иннервирующие мышцы глазных яблок.
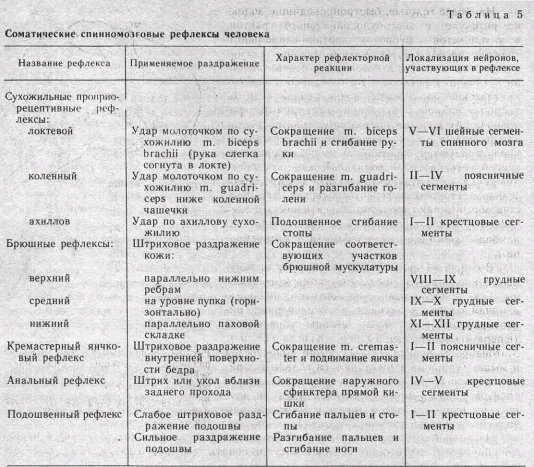
Шейные тонические рефлексы возникают при поворотах и наклонах го'ловы, что вызывает растяжение мышц шеи и активирует рецептивное поле рефлекса. Рефлексы, которые обычно исследуют у человека, приведены в табл. 5.
Кроме рассмотренных выше рефлексов, которые относятся к категории соматических, так как выражаются в активации скелетных мышц, спинной мозг играет важную роль в рефлекторной регуляции внутренних органов, являясь центром многих висцераль-
ны.х рефлексов. Эти рефлексы осуществляются при участии расположенных в боковых и вентральных рогах серого вещества преганглионарных нейронов вегетативной нервной системы. Аксоны этих нервных клеток покидают спинной мозг через передние корешки и заканчиваются на клетках симпатических или парасимпатических вегетативных ганглиев. Ганглионарные нейроны в свою очередь посылают аксоны к клеткам различных внутренних органов, включая гладкие мышцы кишечника, сосудов, мочевого пузыря, к железистым клеткам, сердечной мышце.
Дата добавления: 2016-03-27; просмотров: 3041;