РЕЦЕПТОРНЫЕ (ЧУВСТВИТЕЛЬНЫЕ) НЕРВНЫЕ ОКОНЧАНИЯ
Рецепторные (чувствительные) нервные окончания воспринимают сигналы из внешней среды (экстерорецепторы) и внутренних органов (интерорецепторы). В зависимости от природы раздражения, регистрируемого рецепторами, они подразделяются в соответствии с физиологической классификацией, на механорецепторы, хеморецепторы, терморецепторы и болевые рецепторы (ноцицепторы). В специализированных органах чувств (орган вкуса, обоняния, зрения, равновесия и слуха) имеются особые рецепторные клетки, которые воспринимают соответствующие раздражения.
Морфологическая классификация чувствительных нервных окончанийоснована на особенностях их структурной организации. В соответствии с этой классификацией различают свободные и несвободные чувствительные нервные окончания; последние включают инкапсулированные и неинкапсулированные окончания (рис. 8.18).
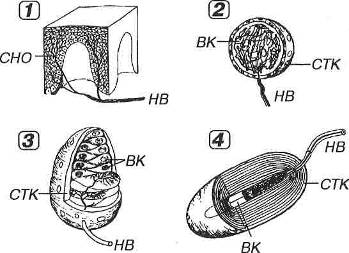
Рис. 8.18. Рецепторные (чувствительные) нервные окончания (по Rohen J.W., Lutjen-Drecoll E. 1982, с изменениями). 1 - свободные нервные окончания (СНО) образованы терминальными ветвлениями дендрита чувствительного нейрона, идущего в составе нервного волокна (НВ). Несвободные инкапсулированные нервные окончания (2-4) образованы ветвлениями дендрита, окруженными леммоцитами, в совокупности с которыми они образуют структуру, называемую внутренней колбой (ВК). Снаружи окончания покрыты соединительнотканной капсулой (СТК). 2 - колба Краузе, 3 - осязательное тельце (Мейснера), 4 - пластинчатое тельце (Фатер-Пачини).
Свободные чувствительные нервные окончаниясостоят только из терминальных ветвлений дендрита чувствительного нейрона, Они встречаются в эпителии, а также в соединительной ткани. Проникая в эпителиальный пласт, нервные волокна утрачивают миелиновую оболочку и нейролемму, а базальная мембрана их леммоцитов сливается с эпителиальной. Свободные нервные окончания обеспечивают восприятие температурных (тепловых и холодовых), механических и болевых сигналов (рис. 8.19).
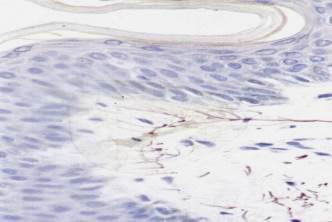
Рис. 8.19. Свободные нервные окончания в соединительной ткани, представленные концевыми ветвлениями дендритов чувствительных нейронов.
Несвободные чувствительные нервные окончаний содержат все компоненты нервного волокна. Они разделяются на инкапсулированные (имеющие особую соединительнотканную капсулу) и неинкапсулированные.
Несвободные неинкапсулированные нервные окончания состоят из ветвлений дендритов, окруженных леммоцитами. Они встречаются в соединительной ткани кожи (дерме), а также собственной пластинки слизистых оболочек.
Несвободные инкапсулированные нервные окончания весьма разнообразны, но имеют единый общий план строения: их основу составляют ветвления дендрита, которые непосредственно окружены леммоцитами и снаружи покрыты особой соединительнотканной капсулой (см. рис. 8.18.). К этому виду нервных окончаний относят пластинчатые тельца (Фатер-Пачини), осязательные тельца (Мейснера) тельца Руффини, колбы Краузе, нервно-мышечные веретена и нервно-сухожильные веретена (сухожильные органы Гольджи).
Пластинчатые тельца (Фатер-Пачини) встречаются в соединительной ткани внутренних органов и кожи (рис. 8.20.). Они имеют вид округлых образований диаметром 1-5мм, воспринимают давление и вибрацию. Структурными компонентами тельца являются:
1) внутренняя колба (луковица), образованная видоизмененными уплощенными леммоцитами, в которую проникают одно или несколько нервных волокон, имеющих прямой ход;
2) наружная колба - слоистая соединительнотканная капсула, состоящая из фибробластов и коллагеновых волокон, образующих 10-60 концентрических пластин, между которыми имеется жидкость.
При деформации пластин капсулы давление передается на нервное окончание, что вызывает деполяризацию его мембраны.
 А А
|  Б Б
|
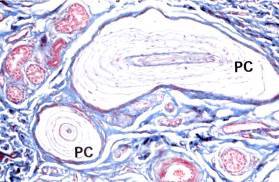
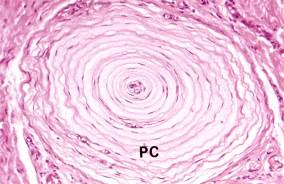
Рис. 8.20. На электронной микрофотографии изображены: Тельца Фатер-Пачини (РС) – инкапсулированный рецептор. Капсула состоит из параллельно расположенной пластинки (по-видимому, производной Шванновской клетки), коллагеновых волокон. В центре капсулы раположены ветвления одного немиелинизированного нервного волокна (х100).
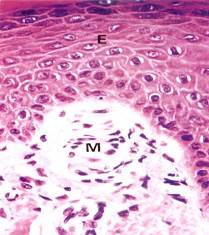
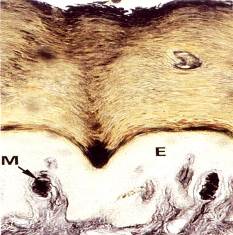
Осязательные тельца (Мейснера) расположены преимущественно в сосочковом слое дермы, имеют эллипсоидную форму и небольшие размеры (около 50-120 мкм) (рис. 8.21.). Их внутренняя колба состоит из плоских глиальных клеток, лежащих перпендикулярно длинной оси тельца, между которыми располагаются веточки дендритов. Между глиальными клетками проникают коллагеновые фибриллы, связанные с базальным слоем эпителия. Капсула тонкая, переходит в периневрий.
 А А
|  Б Б
|
Рис. 8.21. Тельца Мейснера. Инкапсулированный рецептор (М), находящийся в коже губ, гениталий и т.д. Имеет овальную форму, располагается в коже прямо под эпидермисом (Е). Рецептор состоит из капсулы, образованной коллагеновыми волокнами, Шванновскими клетками (х320). (Б метод импрегнации х150). В капсуле находятся несколько безмякотных нервных окончаний, которые обильно ветвятся.).
Тельца Руффини лежат в соединительнотканной части кожи и капсулах суставов; они воспринимают давление и имеют вид веретеновидных структур длиной до 1-2 мм. Внутреннюю колбу образуют глиальные клетки, между которыми располагаются многочисленные ветвящиеся терминали дендритов с расширениями на концах. Капсула хорошо выражена, образована коллагеновыми волокнами.
Колбы Краузе - мелкие (40-150 мкм) округлые тельца, являющиеся механорецепторами и, возможно, холодовыми рецепторами. Они расположены в соединительной ткани сосочкового слоя дермы и собственной пластинке слизистой оболочки полости рта, надгортанника, в конъюнктиве глаза. Внутренняя колба образована уплощенными глиальными клетками, между которыми тонкие веточки дендрита образуют сплетение в виде клубочка. Капсула состоит из плоских клеток, являющихся продолжением периневрия.
Нервно-мышечные веретена - рецепторы растяжения волокон поперечнополосатых мышц - сложные инкапсулированные нервные окончания, обладающие как чувствительной, так и двигательной иннервацией. Число веретен в мышце зависит от ее функции и тем выше, чем более точными движениями она обладает. Нервно-мышечное веретено (рис. 8.22, 8.23.) имеет длину 0,5-7 мм и располагается параллельно ходу волокон мышцы, называемых экстрафузальными (от лат. extra - вне и fuso - веретено, т.е. расположенными за пределами веретена). Веретено покрыто тонкой соединительнотканной капсулой (продолжением периневрия), внутри которой находятся тонкие поперечнополосатые интрафузальные мышечные волокна двух видов:
- волокна с ядерной сумкой - в расширенной центральной части которых содержатся скопления ядер (1-4 волокна/веретено);
- волокна с ядерной цепочкой - более тонкие с расположением ядер в виде цепочки в центральной части (до 10 волокон/веретено).
Чувствительные нервные волокна образуют кольцеспиральные окончания на центральной части ишрафузальных волокон обоих типов и гроздьевидные окончания у краев волокон с ядерной цепочкой.
Двигательные нервные волокна - тонкие, образуют мелкие нервно-мышечные синапсы по краям интрафузальных волокон, обеспечивая их тонус.
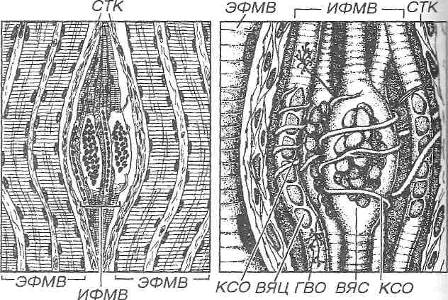
Рис. 8.22. Нервно-мышечное веретено. 1 - общий вид веретена, располагающегося между экстрафузальными мышечными волокнами (ЭФМЗ) и образованного интрафузальными мышечными волокнами (ИФМВ), которые окружены соединительнотканной капсулой (СТК). 2 - детали строения веретена, содержащего два вида ИФМВ: волокна с ядерной сумкой (ВЯС) и волокна с ядерной цепочкой (ВЯЦ). Чувствительные нервные волокна образуют кольцеспиральные окончания (КСО) на центральной части ИФМВ обоих типов и гроздьевидные окончания (ГВО) у краев ВЯЦ. Нервно-мышечное веретено содержит также двигательные нервные волокна и образованные ими нервно-мышечные синапсы по краям ИФМВ (не показаны).
 А А
|  Б Б
|
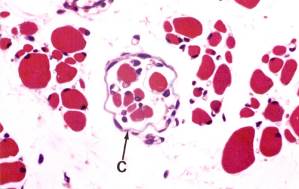
Рис. 8.23. Нервно-мышечное веретено N-нервное волокно, С – капсула.А – продольный срез (х320), Б – поперечный.
Нервно-сухожильные веретена (сухожильные органы Гольджи) - рецепторы растяжения - веретеновидные инкапсулированные структуры длиной около 0.5-1мм, располагающиеся в области соединения волокон поперечнополосатых мышц с коллагеновыми волокнами сухожилий. Каждое веретено образовано капсулой из плоских фиброцитов (продолжение периневрия), которая охватывает группу сухожильных пучков, оплетенных многочисленными терминальными веточками нервных волокон, частично покрытых леммоцитами. Возбуждение рецепторов возникает при растяжении сухожилия во время мышечного сокращения.
Дата добавления: 2016-02-16; просмотров: 1980;