ПРИНЦИП СЕКВЕНИРОВАНИЯ ДНК МЕТОДОМ ХИМИЧЕСКОЙ ДЕГРАДАЦИИ ПО МАКСАМУ-ГИЛБЕРТУ
В основе метода секвенирования ДНК путем химической деградации лежит ограниченное расщепление меченого фрагмента ДНК под действием специфических реагентов. Условием проведения секвенирования этим методом является наличие фрагмента ДНК, меченного только по одному концу. Разделение продуктов деградации по размеру с помощью электрофореза в полиакриламидном геле, способного разделять фрагменты ДНК, различающиеся между собой по длине всего на один нуклеотид в широком диапазоне. Определить нуклеотидную последовательность секвенированного участка ДНК позволяет последующая радиоавтография геля.
Сначала проводится ограниченная модификация определенных нуклеотидов под действием различных химических агентов. Концентрация агента и продолжительность его воздействия на молекулы ДНК подбирается с расчетом, чтобы в каждой молекуле произошла модификация только одного нуклеотида, а поскольку в реакционной смеси присутствует огромное количество таких молекул, то, согласно теории вероятности, все основания данного типа в секвенируемом фрагменте ДНК окажутся модифицированными. Следующие этапы удаления модифицированных оснований и р-элиминации обоих фосфатов, окружающих дезоксирибозу, и разрыва цепи должны проходить уже количественно. Отдельные реакции ограниченной модификации и количественного расщепления проводят для каждого типа нуклеотидов или их комбинации. Таким образом, в результате четырех (или иногда трех, пяти или даже шести) типов реакций образуется смесь олигонуклеотидных молекул, которые различаются по размеру на один нуклеотид и несут на одном из концов метку, обычно радиоактивную. Также кроме меченых молекул в реакционной смеси будут находится, и фрагменты, не несущие метки, но они окажутся невидимыми на этапе радиоавтографии. Восстановить последовательность нуклеотидов секвенируемого фрагмента ДНК можно после разделения продуктов реакции в дорожках денатурирующего полиакриламидного геля и этапа радиоавтографии. На рентгеновской пленке будет видна лестница из полос ДНК.
На рисунке 1 в виде схемы показан процесс определения последовательности нуклеотидов ДНК данным методом.
ПРИНЦИП СЕКВЕНИРОВАНИЯ ДНК ФЕРМЕНТАТИВНЫМ МЕТОДОМ
ПО СЭНГЕРУ
В основе этого метода секвенирования (также называемого секвенированием путем терминации цепи), лежит ферментативное построение комплементарной цепи ДНК на одноцепочечной матрице. При этом в разных местах цепи ДНК происходит ингибировании ее дальнейшего роста.
Компонентами реакции являются: одноцепочечная матрица ДНК; короткая затравочная молекула, комплементарная определенному участку матрицы; ДНК-полимераза (Кленовский фрагмент ДНК-полимеразы I E. coli); 2´-дезоксинуклеотид 5´-трифосфаты (дАТФ, дГТФ, дТТФ, дЦТФ или просто дНТФ); 2´,3´-дидезоксинуклеотид 5´-трифосфаты (ддАТФ, ддГТФ, ддТТФ, ддЦТФ или просто ддНТФ); реакционный буфер с ионами Mg2+.
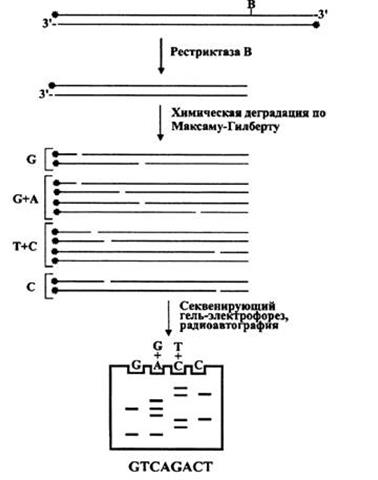
Рисунок 1 – Принцип секвенирования методом Максама-Гилберта
Главным этапом этого процесса является терминация построения комплементарной цепи ДНК. Она происходит при включении ДНК-полимеразой модифицированных аналогов природных субстратов дидезоксинуклеотид трифосфатов, являющихся ингибиторами ДНК-полимеразы. Рост цепи таким образом прерывается из-за отсутствие второго гидроксильного остатка в З'-положении рибозного кольца у дидезоксипроизводных и невозможности присоединения к ним следующего нуклеотида. Поскольку в четырех реакционных пробирках (по типу оснований – А, С, G и Т) присутствует огромное количество молекул секвенируемой ДНК, во много раз превосходящее ту длину фрагмента, которую могла построить ДНК-полимераза, то по теории вероятности каждое положение этого типа оснований во фрагменте ДНК оказывается представленным соответствующим ддНМФ.
Еще одним важным моментом является разный размер полученных фрагментов ДНК. Разница в размерах определяется тем, что все З'-концы фрагментов вновь синтезируемой цепи ДНК во всех четырех реакционных пробирках были терминированы соответствующими ддНМФ и, таким образом, представляли собой полный набор всех возможных длин в пределах секвенируемого участка, построенного ДНК-полимеразой. Что касается 5'-конца всех этих фрагментов ДНК, принадлежащих новой цепи, то он должен быть для всех фрагментов строго одинаковым и принадлежать 5'-концу исходной затравочной молекулы.
Продукты реакций терминирования подвергаются денатурации и одноцепочечные меченые фрагменты разделяются по длине посредством электрофореза в полиакриламидном геле, позволяющим разделять фрагменты ДНК, отличающиеся всего на один нуклеотид.
После электрофореза гель экспонировался на рентгеновскую пленку. С проявленной пленки “читали” последовательность нуклеотидов секвенируемого участка ДНК.
На рисунке 2 схематично показан описанный выше процесс.
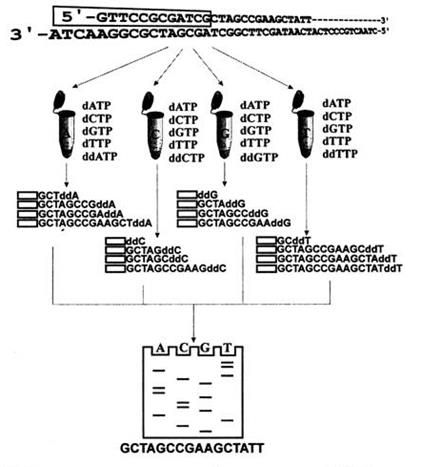
Рисунок 2 – Принцип секвенирования методом Сенгера
Дата добавления: 2016-02-09; просмотров: 4257;