Рост популяций и кривые роста
Если рождаемость в популяции превышает смертность, то популяция, как правило, будет расти. С увеличением плотности скорость роста популяции постепенно снижается до нуля. При нулевом росте популяция стабильна, т. е. размеры ее не меняются. Отдельные организмы при этом могут расти и размножаться. Нулевая скорость роста означает лишь то, что скорость размножения, если оно происходит, уравновешена смертностью. Данная картина характерна для ряда одноклеточных и многоклеточных организмов, например для клеток водорослей в культураль-ной жидкости, для фитопланктона озер и океанов весной, для насекомых (мучные хрущаки, а также клещи, интродуцирован-ные в новое местообитание с обильными запасами пищи, где нет хищников).
Миграция, или расселение, так же как и внезапное снижение скорости размножения, может способствовать уменьшению численности популяции. Расселение может быть связано с определенной стадией жизненного цикла, например с образованием семян. Рассматривая вопрос об оптимальных размерах популяции в данной среде, следует учитывать поддерживающую емкость или кормовую продуктивность среды. Чем выше поддерживающая емкость, тем больше максимальный размер популяции, который может существовать неопределенно долгое время в данном местообитании. Дальнейшему росту популяции будут препятствовать один или несколько лимитирующих факторов. Это зависит от доступности ресурсов для данного вида.
Таким образом, скорость роста популяции в естественных местообитаниях будет зависеть от климатических изменений, от снабжения пищей и от того, ограничено ли размножение определенным временем года и др., что должно учитываться при составлении моделей или их усовершенствовании.
Математические модели экспоненциального роста популяций и роста при ограниченных ресурсах. Рост численности популяции в геометрической прогрессии можно описать с помощью простых уравнений. Так, в популяции с исходной численностью в N особей за промежуток времени Dt появляется DN новых особей. Если число вновь появившихся особей прямо пропорционально N и Dt, то имеем уравнение DN = r ×Dt ×N. Разделив обе его части на Dt, получим
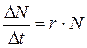 (9.4)
(9.4)
Величина 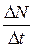 - абсолютная скорость роста численности,
- абсолютная скорость роста численности,
г — биотический потенциал или удельная скорость роста численности.
За малый промежуток времени изменение численности равно ее производной 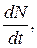 и уравнение (9.4) можно переписать так:
и уравнение (9.4) можно переписать так:
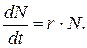 (9.5)
(9.5)
Решение этого уравнения — функция
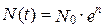 . (9.6)
. (9.6)
Здесь е — основание натуральных логарифмов (е » 2,72...). График этой функции и есть экспонента (рис. 9.10,вверху).
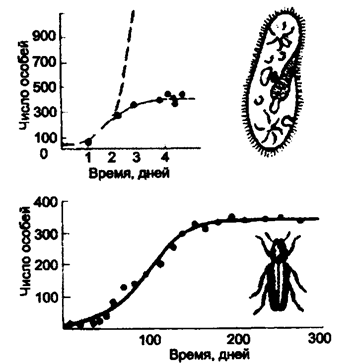
Рис. 9.10. Реальная и теоретическая кривые роста численности инфузорий-туфелек (вверху) и рост численности жуков определенного вида в культуре (численность меняется по правилам логистического роста)
Пунктирная линия — теоретическая кривая (экспонента); сплошная линия — в реальной культуре рост численности замедляется и через определенное время останавливается
В модели экспоненциального роста удельную рождаемость b и удельную смертность d можно обозначить как 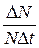 .
.
При этом в замкнутой популяции
DN » b×N×Dt - d×N×Dt;
r = b – d. (9.7)
Если смертность выше рождаемости, то убывание численности тоже описывается уравнением (9.6), но с отрицательным г. Такой процесс называют экспоненциальным затуханием численности.
Модель динамики численности популяции при органиченных ресурсх предложил в 1845 г. французский математик Ферхюльст. Уравнение, которое носит его имя, выглядит так:
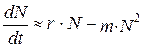 (9.8)
(9.8)
Уравнение Ферхюльста отличается от уравнения экспоненциального роста тем, что в правой его части добавляется выражение mN2. Это выражение учитывает число встреч животных, при которых они могут конкурировать за какой-либо ресурс: вероятность встречи двух особей пропорциональна квадрату численности (точнее, плотности) популяции. У многих животных рост численности популяции действительно ограничивается именно частотой встреч особей.
Перепишем уравнение Ферхюльста следующим образом:
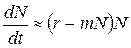 (9.9)
(9.9)
Выражение в скобках — удельная скорость роста численности. Здесь она непостоянна и убывает с увеличением численности популяции. Это отражает усиление конкуренции за ресурсы по мере роста численности.
Если в уравнении (5) вынести в правой части rN за скобки и обозначить 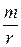 за
за 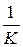 , то получим:
, то получим:
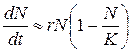 (9.10)
(9.10)
Если N мало по сравнению с k, то выражение в скобках близко к единице: при этом уравнение (9.10) переходит в уравнение экспоненциального роста. График роста численности будет при малых N близок к экспоненте. Когда N близко к k, выражение в скобках близко к нулю, т. е. численность популяции перестает увеличиваться. Отсюда ясно, что k в данной модели — это и есть емкость среды. При N больших, чем k, абсолютный прирост численности становится отрицательным, и численность убывает до величины, равной емкости среды. График зависимости численности популяции от времени, соответствующий решению уравнения (9.10), — S-образная кривая, подобная изображенной на рис. 9.10, внизу. Эта кривая называется логистической кривой, а рост численности, соответствующий уравнению (9.10), —логистический рост.
На логистической кривой есть точка, где абсолютная скорость роста численности максимальна. Можно показать, что максимальная скорость роста достигается, когда численность равна.
Популяции, существующие в условиях ограниченных ресурсов, нередко хорошо подчиняются правилам логистического роста.
Дата добавления: 2015-12-01; просмотров: 1099;