Произвольные движения и их патология
Активные движения человека, которые формируются в процессе жизненного опыта и непрерывно контролируются всей деятельностью мозга, называют произвольными, или сознательными. Двигательный акт является сложным рефлексом, который осуществляется при участии различных уровней нервной системы. Контролирующая роль в формировании движений принадлежит, главным образом, афферентным сигналам. Прежде чем возникнет запланированное движение, сенсорные системы коры большого мозга по чувствительным проводящим путям получают информацию об исходном состоянии суставно-мышечного аппарата, о готовности его к выполнению движения. Афферентные импульсы от проприорецепторов постоянно сообщают о необходимости осуществления того или иного произвольного движения, о ходе его выполнения, осуществляют своевременную коррекцию силы, длительности, последовательности мышечных сокращений, информируют о целесообразности приостановления движения или о его завершении. Чувствительная и двигательная зоны коры образуют единую сенсомоторную систему, регулирующую работу нижерасположенных нервных центров и осуществляет регуляцию двигательной активности человека. Действие проприоцептивного анализатора дополняется функционированием вестибулярного, зрительного, слухового анализаторов. Важную роль в создании точных, координированных движений играют многочисленные образования экстрапирамидной системы, мозжечок, промежуточный мозг.
Двигательные центры коры большого мозга расположены преимущественно спереди центральной борозды, т.е. в лобной доле мозга. Это предцентральная извилина, прицентральная долька и прилегающие участки лобной доли. В передних отделах лобных долей полушарий мозга (третичное проекционное поле) происходит интеграция всех информационных сигналов, формируется модель двигательного акта. Произвольные движения осуществляются также при участии премоторного участка (вторичные проекционные двигательные поля). Реализация плана движения непосредственно осуществляется предцентральной извилиной и прицентральной долькой коры. Здесь в пятом слое коры большого мозга расположены гигантские пирамидные двигательные клетки, описанные в 1874р. киевским нейрогистологом В.А. Бецом. Аксоны клеток Беца образуют пирамидный путь, который вместе с двигательными волокнами периферической нервной системы и мышечным аппаратом обеспечивают выполнение заданной двигательной программы.
Раздражение тел мотонейронов в предцентральной извилине вызывает приступы клонических или тонических судорог, которые называют джексоновской эпилепсией в честь английского ученого Jackson John Hughlings (1835-1911), который их описал. Характерным для эпилепсии джексоновского типа является начало судорог на ограниченной группе мышц при ясном сознании больного. Иногда судороги генерализуються и переходят в общий приступ, при котором больной теряет сознание.
Раздражение отдельных участков предцентральной извилины вызывает фокальные судороги в соответствующих группах мышц с противоположной стороны, поскольку каждый из отделов этой зоны коры связан с определенным участком тела. В прицентральной дольке и в верхней четверти предцентральной извилины расположены центры для иннервации мышц нижней конечности (сверху - стопа, ниже - голень и бедро), в средних двух четвертях извилины центры для иннервации мышц верхней конечности (сверху - плеча, предплечья, внизу - кисти), в нижней четверти извилины проецируются мышцы лица, глотки гортани, языка.
Проекция мышц головы (включая мышцы глаза) локализована в заднем отделе средней лобной извилины, а мышц туловища - в заднем отделе верхней лобной извилины (соматотопическое представительство двигательных функций). Площадь каждого из двигательных участков зависит не от массы мышц, а от сложности и точности функций, ими выполняемых. Наибольшее представительство в коре имеют мышцы, которые выполняют самые сложные дифференцированные движения (кистей, пальцев рук, губ, языка). Для осуществления произвольного движения импульсы, возникшие в коре большого мозга, должны поступить в соответствующие мышцы. Это обеспечивается при помощи пути, который клиницисты называют корково-мышечным (рис. 1).
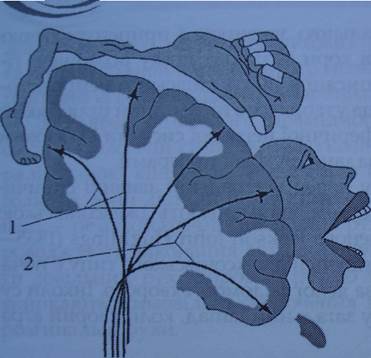
Рис. 1. Соматотопическое представительство в двигательной зоне коры большого мозга: 1. - Корково-спинномозговой путь; 2 - корково-ядерный путь
Этот путь составляют два нейрона - центральный и периферический. Центральный нейрон - это тела гигантских пирамидных клеток (Беца) с их аксонами, периферический - тела и аксоны клеток передних рогов спинного мозга, двигательные ядра и корешки черепных нервов. По аксонам пирамидных клеток импульсы идут от коры большого мозга к мотонейронам передних рогов спинного мозга и двигательных ядер черепных нервов, образуя пирамидный путь. Поэтому пирамидный путь состоит из двух пучков нервных волокон - корково-спинномозговых (от коры до спинного мозга) и корково-ядерных (от коры к двигательным ядрам черепных нервов). Пирамидные пути размещаются под корой в виде лучистого венца, а дальше, постепенно сближаясь друг с другом, проходят между подкорковыми ядрами, образуя колено и передние две трети задней ножки внутренней капсулы. Далее волокна пирамидного пути идут через основание ножек мозга моста и продолговатого мозга, на передней поверхности которого они образуют два заметных валика - пирамиды.
На границе между продолговатым мозгом и спинным, корково-спинномозговые волокна частично перекрещиваются. Большая, перекрещенная часть волокон переходит в боковые канатики спинного мозга (боковой перекрещенный корково-спинномозговой (пирамидный) путь), меньшая, неперекрещеная часть переходит в передние канатики спинного мозга (передний неперекрещений корково-спинномозговой (пирамидный) путь). Пучки волокон корково-спинномозгового пути, постепенно истончаясь, заканчиваются на альфа-мотонейронах передних рогов спинного мозга всех сегментов, которым они передают двигательные импульсы (рис. 2). Волокна переднего корково-спинномозгового пути переходят на противоположную сторону на уровне сегментов в составе передней белой спайки спинного мозга. Предполагают, что в шейных и грудных сегментах спинного мозга некоторые из этих волокон соединяются с клетками переднего рога своей стороны, благодаря этому мышцы шеи и туловища получают корковую иннервацию с обеих сторон. Аксоны периферических нейронов сначала выходят в составе передних корешков спинного мозга, далее в составе нервных сплетений и периферических нервов направляются в скелетные мышцы.

Рис. 2. Схема хода корково-спинномозгового и корково-ядерного путей:
1 - предцентральная извилина, 2 - ножка мозга, 3 - мост, 4 - продолговатый мозг; 5 - перекресток пирамид; 6 - шейное утолщение спинного мозга; 7 - поясничное утолщение спинного мозга
Волокна корково-ядерного пути, заканчивающиеся на двигательных ядрах черепных нервов мозгового ствола, передают импульсы на моторные клетки этих ядер, их аксоны образуют двигательные волокна черепных нервов, иннервирующие соответствующие мышцы. Волокна корково-ядерного пути также осуществляют частичное пересечение над ядрами черепных нервов (см. рис. 2). Только волокна, идущие к нижней части ядра лицевого нерва и до ядра подъязычного нерва, осуществляют полный надъядерный перекрест. Благодаря неполному перекресту пирамидного пути, обеспечивается двусторонняя корковая иннервация мышц. Но она не во всех группах мышц одинакова. Больше всего она выражена в мышцах, иннервируемых черепными нервами, в мышцах шеи, туловища, промежности, меньше в мимических мышцах нижней части лица, мышцах языка и конечностей. Именно эти мышцы имеют преимущественно одностороннюю корковую двигательную иннервацию из противоположного полушария головного мозга. Поэтому в случае одностороннего поражения пирамидного пути теряются движения с противоположной стороны именно в мышцах конечностей, языка и нижней половины лица.
Функция большинства мышц, иннервируемых черепными нервами, мышц шеи, туловища и промежности не нарушается. Таким образом, первые нейроны двигательного пути осуществляют связь двигательной области коры с сегментарным аппаратом спинного мозга и мозгового ствола. Весь комплекс клеток, с помощью которых осуществляется эта связь, называется пирамидной системой. Функции пирамидной системы: принимает участие в выполнении произвольных движений, посылая импульсы к стволовым и спинальным сегментарным аппаратам; регулирует функции сегментарных аппаратов, активирует большие альфа-мотонейроны передних рогов спинного мозга и тормозит деятельность рефлекторных дуг; ограничивает распространение импульсов возбуждения по нейронам спинного мозга; тормозит рефлекторные автоматизмы подкоркового, стволового и спинального уровней.
Если двигательный корково-мышечный путь поражается на любом уровне, то соответствующие мышцы не получают иннервации от коры, произвольные движения в мышцах становятся невозможными, мышца перестает сокращаться, возникает ее паралич (плегия). В случае частичного поражения двигательного пути наблюдается парез - неполная потеря произвольных движений, ограничения их объема, обусловленные нарушением иннервации мышц. Поскольку гигантские пирамидные клетки с их длинными аксонами, которые образуют пирамидный путь, являются центральными нейронами, то паралич или парез мышц, возникающий вследствие их повреждения, называется центральным. Мотонейроны передних рогов спинного мозга и двигательных ядер черепных нервов называют периферическими, поэтому паралич, возникающий в случае их поражения, называют периферическим. Следовательно, вид паралича определяется названием пораженного нейрона.
Особенности центрального паралича (пареза):
1. Поражение центральных нейронов охватывает целые пучки волокон пирамидного пути. Поэтому обычно возникает центральный паралич не отдельных мышц, а целых их групп
2. Центральный паралич (парез) носит название спастического, поскольку сопровождается повышением мышечного тонуса. Вследствие поражения центральных нейронов снимается контроль пирамидной системы за тонической деятельностью сегментарного аппарата, за функцией спинальных рефлекторных дуг. Преимущественно мышечный тонус усиливается в разгибателях нижней конечности и сгибателях верхней. Это способствует возникновению характерной позы Вернике-Манна (особенно вследствие поражения внутренней капсулы) - парализованная верхняя конечность приведена к туловищу, пронирована и согнута в локтевом суставе, кисть и пальцы также согнуты, а нижняя конечность разогнута в тазобедренном и коленном суставах, будто вытянутая и «удлиненная ». Во время шествия больной этой ногой описывает полукруг, чтобы не задевать пола носком разогнутой ноги.
3. Повышаются сухожильные и периостальные рефлексы. Это объясняется тем, что растормаживаются рефлекторные дуги на уровне сегментарного аппарата. При таких условиях вследствие значительной сухожильной гиперрефлексии возникает клонус стопы, кисти.
4. Подавляются кожные брюшные, подошвенные рефлексы, возникновение которых связано с формированием пирамидных путей.
5. Появляются патологические рефлексы: появление подошвенных сгибательных или разгибательных патологических признаков свидетельствует о поражении корково-спинномозговых волокон, а рефлексов орального автоматизма - о двустороннем поражении корково-ядерных путей.
6. Вследствие растормаживания сегментарно-рефлекторного аппарата усиливаются спинальные автоматизмы (защитные движения), появляются патологические синкинезии.
Центральный паралич мышц нередко сопровождают расстройства мочеиспускания и дефекации. Центры этих функций расположены в сером веществе спинного мозга на уровне боковых рогов S2-S4 сегментов. Сознательный контроль за мочеиспусканием обеспечивается благодаря связям этих центров с корой полушарий головного мозга. Корковая иннервация тазовых органов осуществляется волокнами пирамидных путей, их двустороннее поражение сопровождается расстройствами этих функций. Возникает периодическое недержание мочи (периодическое рефлекторное опорожнение мочевого пузыря без сознательного контроля в случае растяжения его мочой), иногда наблюдается задержка мочи, императивные позывы к мочеиспусканию.
Особенности периферического паралича (пареза)
1. Поражения периферического нейрона в любом его участке приводит не только к разрыву двигательного корково-мышечного пути, но и к разрыву сегментарной рефлекторной дуги в ее эфферентном звене. Поэтому при наличии периферического паралича становятся невозможными как произвольные, так и рефлекторные движения. Возникает арефлексия или гипорефлексия рефлексов, замыкающихся на уровне поражения.
2. Наблюдается мышечная атония или гипотония, поскольку вследствие разрыва спинальной рефлекторной дуги мышечные волокна не получают тонических импульсов, поэтому периферический паралич получил название вялого.
3. Через 2-3 недели после начала повреждения периферических нейронов в соответствующих мышцах наблюдается атрофия, потому что к ним вследствие разрыва рефлекторной дуги не поступают трофические влияния.
4. Три характерных признака периферического паралича: арефлексия, атония и атрофия мышц могут дополняться другими симптомами. В случае хронических прогрессирующих процессов в передних рогах спинного мозга или в двигательных ядрах черепных нервов возникает раздражение тел периферических мотонейронов и наблюдаются фибриллярные подергивания мышц.
5. Распространение периферического паралича в основном ограничено, поскольку поражаются преимущественно отдельные участки передних рогов, отдельные передние корешки или периферические нервы.
6. Периферический паралич характеризуется возникновением реакции перерождения или дегенерации, которая проявляется изменениями реакций на электрический ток пораженных нервов и мышц, которыми иннервируются.
Периферические параличи возникают при наличии поражения передних рогов спинного мозга, двигательных ядер черепных нервов и их корешков, передних корешков спинного мозга, шейного, плечевого и пояснично-крестцового сплетений, периферических нервов.
Дата добавления: 2015-12-22; просмотров: 1205;