Скелетная мускулатура
Функциональная организация скелетной мускулатуры - двигательные единицы (ДЕ)
• ДЕ – один мотонейрон и группа иннервируемых им мышечных волокон.
• Чем меньше волокон в одной ДЕ, тем более тонкое движение можно совершить.
• По лабильности ДЕ:
• быстрые (длительность ПД от 1 мс, высокая скорость сокращения, гидролиза и ресинтеза АТФ)
• медленные (длительность ПД до 10-15 мс, низкая скорость сокращения, гидролиза и ресинтеза АТФ).
Механизм мышечного сокращения
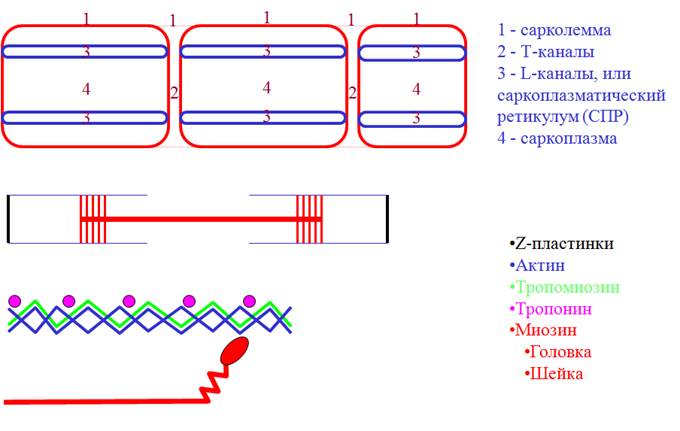
• Распространение ПД по Т-каналам ® деполяризация СПР ® выход Са2+.
• Са2+-тропонин ® сдвиг тропомиозина ® открытие центров связывания головки миозина на актине.
• Соединение головки миозина с актином ® жесткость (ригидность) мышцы.
• АТФ-азная активность головки миозина ® гидролиз АТФ ® гребковое движение (генерация усилия).
• Присоединение новой молекулы АТФ к миозиновой головке ® разрыв поперечного мостика (исчезновение ригидности).
• Присоединение головки миозина к другому центру актина.
• Активация Са-АТФазы СПР фосфатом (гидролиз АТФ) ® возврат Са2+ обратно ® расслабление мышцы.
Регуляция силы и скорости сокращения
Суммация
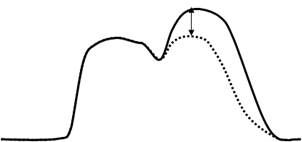
• амплитуды сокращения – после первого сокращения часть Са2+ не успевает возвратиться в СПР (движение против градиента концентраций требует АТФ) и суммируется с вновь поступившей порцией.
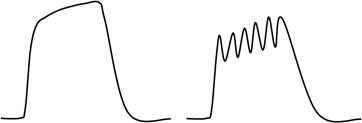
• Слитное сокращение – тетанус.
– Гладкий – новая фаза укорочения накладывается на предыдущую.
– Зубчатый – новая фаза укорочения накладывается на фазу удлинения.
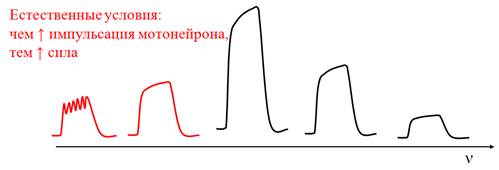
• В естественных условиях гладкий тетанус целой мышцы возникает за счет несинхронности зубчатых тетанусов отдельных ДЕ.
• Так как возврат Са2+ медленнее его выхода, чем частота возбуждения, тем амплитуда и сила сокращения;
• Но небезгранично: имеется оптимум частоты возбуждения, после которого развивается пессимальное торможение (пессимум).
Зависимость длина-сила (зависимость Франка-Старлинга)
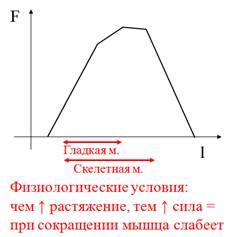
Зависимость длина-сила (зависимость Франка-Старлинга)-зависимость силы мышцы от степени ее растяжения
¯ силы при больших длинах:
• ¯ перекрывания нитей актина и миозина
¯ силы при малых длинах:
• ¯электромеханического сопряжения;
• “сморщивание” саркомера
Режимы сокращений
• Изотонический – при постоянном напряжении
• Изометрический – при постоянной длине (без укорочения) – максимальная сила
• Концентрический – усилие при укорочении
• Эксцентрический – усилие при удлинении
Зависимость нагрузка-скорость (сила-скорость)
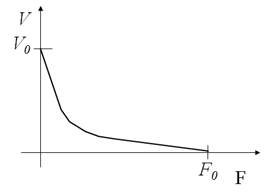
Чем больше нагрузка, тем медленнее сокращение. Чем быстрее сокращение, тем оно слабее.
Причины:
• Длительность актомиозинового взаимодействия ~ сила.
• При большой скорости укорочения поперечные мостики не успевают распадаться.
• F0 (изометрическая сила) зависит от количества поперечных мостиков (определяется длиной саркомера и количеством вышедшего Са2+).
• V0 (cкорость при нулевой нагрузке) зависит от скорости оборота поперечных мостиков и гидролиза АТФ (определяется типом волокон).
Последовательное и параллельное соединение саркомеров (волокон)
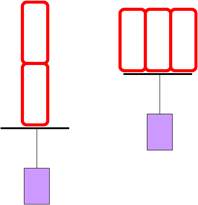
• Параллельное соединение (толстые мышцы) увеличивает силу
• Последовательное соединение (длинные мышцы) - скорость
Регуляция силы и скорости мышечных сокращений – выводы:
Скорость сокращениязависит от
• скорости оборота поперечных мостиков и активности АТФазы миозина
• нагрузки – зависимость нагрузка-скорость
• числа активных ДЕ (при числа ДЕ нагрузка на одно волокно ¯) – зависимость нагрузка-скорость
• анатомической длины мышцы (числа последовательно соединенных саркомеров)
Сила сокращениязависит от
• степени растяжения мышцы – зависимость длина-сила
• скорости укорочения – зависимость нагрузка-скорость
• частоты импульсации мотонейрона – суммация сокращений
• числа активных ДЕ – нагрузка на одно волокно
• толщины мышц (волокон).
Дата добавления: 2015-11-06; просмотров: 1070;