Круги Пейпса и Наута
Функции лимбической системы мозга, состоящей из кругов Пейпса и Наута, в психических явлениях человека и высших животных и являются предметом познания психофизиологии. Психофизиология и нейропсихология - вот две базовые теории в построении общей психологии.
Важной структурой круга Пейпса является гиппокамп, благодаря которому поддерживается информация о текущих событиях. По орбитам круга Пейпса циркулируют паттерны - коды информации из нервных импульсов. Эти паттерны через таламус и поясную извилину передаются всей коре больших полушарий.
Круг Наута проходит через гипоталамус, через который психика управляет всей гормональной активностью организма. Круг Пейпса оперирует информацией, в круге Наута формируются мотивы ответного поведения. Оба круга взаимосвязаны через связь гипоталамуса с мамиллярным телом.
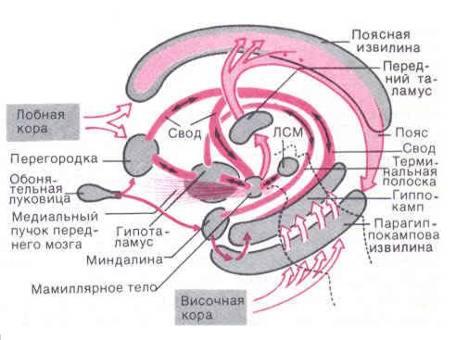
Круг Пейпса.Нейрофизиологам известна лимбическая система мозга (архипалеокортекс) под новой корой больших полушарий (неокортекс). Эту систему ещё называют эмоциональным мозгом - её мы и возьмём за основу для объяснения психических явлений человека.
Лимбическая система состоит из огромного множества замкнутых нервных волокон в которых циркулируют (реверберируют) нервные импульсы. Назовём одно такое волокно (замкнутую цепь нейронных аксонов) орбитой, систему нервных импульсов в орбите с определённым информационным кодом паттерном.
 Круг Пейпса лимбической системы состоит из функционально важных для психики человека структур: гиппокамп, гипоталамус, передний таламус, поясная извилина.
Круг Пейпса лимбической системы состоит из функционально важных для психики человека структур: гиппокамп, гипоталамус, передний таламус, поясная извилина.
Поясная извилина имеет связь со всей корой больших полушарий. Гиппокамп является информационным центром лимбической системы . Гиппокамп имеет два вида нейронов, одни из которых поддерживают в памяти незнакомую информацию в течение 30 дней. Паттерн ы незнакомой информации от гиппокампа идут к гипоталамусу, повышая его гормональную активность и эмоциональное состояние, но в таламусе они прерываются. Таламус есть своего рода фильтр, который пропускает только известную информацию и отправляет её в поясную извилину, в неокортекс для формирования ответного поведения.
Не всегда следует с получением сведения сразу же запускать поведение, необходимо выждать время. И вся информация, отфильтрованная таламусом, реверберирует в круге Пейпса, её поддерживает второй вид нейронов гиппокампа. Паттерны, совершающие полный оборот по кругу Пейпса, есть сознание. Сознание есть вся совокупность оперативной информации, необходимая для регулирования текущего поведения. Как только поведение достигает определённой цели, имеющееся сознание в лимбической системе затухает, человек переходит к новому сознанию.
Гиппокамп, пропуская через себя сознание, одновременно поддерживает и подсознание, то есть те паттерны, которые не пропускает таламус, не укладываются они в его логических связях. Подсознание действует на гипоталамус, обуславливает состояние тревожности, но человек его не осмысливает, не понимает.
Вся совокупность сведений о предмете фиксируется в матрице. Матрица состоит из орбит и паттернов в этих орбитах. Причём, как и в матрице из линейной алгебры, на каждой орбите (строке) определённое количество паттернов (элементов). Достигается это в результате латерального торможения: если орбита более возбуждённая, имеет высокую плотность паттернов, то она доминирует и затормаживает соседние орбиты. Таким образом, в матрице сохраняются орбиты с равными плотностями паттернов. Менее возбуждённые орбиты отражают сведения о предмете, к которым субъективно мало интереса - и они выпадают из матрицы, из сознания.
Выпадение орбит из матрицы назовём редукцией. Редукция бывает эмоциональной и логической. Эмоциональную редукцию осуществляют доминирующие орбиты, затормаживающие орбиты с меньшим возбуждением. Логическая редукция протекает в структуре таламуса посредством нейронов с тормозным медиатором. Если два каких-то сведения исключают третье сведение, то соответствующие им орбиты затормаживают третью орбиту.
Вообще, ядра таламуса, имеющие связь с лимбической системой , назовём рассудком. Рассудок имеет наследственные или приобретённые логические связи из нейронов, которые обрабатывают сведения о предмете и направляют их к определённым нейронам неокортекса.
Функциональной единицей неокортекса является не нейрон, а колонка, состоящая из звёздчатых и одного пирамидного нейрона, расположенных друг над другом в шести слоях коры. У каждой матрицы есть определитель - это колонка неокортекса, соответствующая предмету.
 Матрица реверберирует по кругу Пейпца , находится в сознании и одновременно поддерживает активность определителя. Если матрица не поддерживает активность определителя, то определитель спит и никак не реагирует на предмет восприятия.
Матрица реверберирует по кругу Пейпца , находится в сознании и одновременно поддерживает активность определителя. Если матрица не поддерживает активность определителя, то определитель спит и никак не реагирует на предмет восприятия.
Круг Наута в лимбической системе имеет меньший размер , чем круг Пейпца , но функциональное значение его гораздо выше.
Реверберируют по кругу нервные импульсы проприоцепций из рецепторов мышц. Сокращает рецепторы мышц лобная кора неокортекса.
Если таламус есть рассудок, то лобная кора (монадная область) есть разум человека.
Классификация рецепторов.
В основу классификации рецепторов положено несколько критериев.
v Психофизиологический характер ощущения: тепловые, холодовые, болевые и др.
v Природа адекватного раздражителя: механо-, термо-, хемо-, фото-, баро-, осмбрецепторы и др.
v Среда, в которой рецептор воспринимает раздражитель: экстеро-, интерорецепторы.
v Отношение к одной или нескольким модальностям: моно- и полимодальные (мономодальные преобразуют в нервный импульс только один вид раздражителя - световой, температурный и т. д., полимодальные могут несколько раздражителей преобразовать в нервный импульс - механический и температурный, механический и химический и т. д.).
v Способность воспринимать раздражитель, находящийся на расстоянии от рецептора или при непосредственном контакте с ним: контактные и дистантные.
v Уровень чувствительности (порог раздражения): низкопороговые (механорецепторы) и высокопороговые (ноцицепторы).
v Скорость адаптации: быстроадаптирующиеся, (тактильные), медленноадаптирующиеся (болевые) и неадаптирующиеся (вестибулярные рецепторы и проприорецепторы).
v Отношение к различным моментам действия раздражителя: при включении раздражителя, при его выключении, на протяжении всего времени действия раздражителя.
v Морфофункциональная организация и механизм возникновения возбуждения: первичночувствующие и вторичночувствующие.
В первичночувствующих рецепторах стимул действует на воспринимающий субстрат, заложенный в самом сенсорном нейроне, который при этом возбуждается непосредственно (первично) раздражителем. К первичночувствующим рецепторам относятся: обонятельные, тактильные рецепторы и мышечные веретена.
К вторичночувствующим относятся те рецепторы, у которых между действующим стимулом и сенсорным нейроном располагаются дополнительные рецептирующие клетки, при этом сенсорный нейрон возбуждается не непосредственно стимулом, а опосредовано (вторично) - потенциалом рецептирующей клетки. К вторичночувствующим рецепторам относятся: рецепторы слуха, зрения, вкуса, вестибулярные рецепторы.
Механизм возникновения возбуждения у этих рецепторов различен. В первичночувствующем рецепторе транформация энергии раздражителя и возникновение импульсной активности идет в самом сенсорном нейроне. У вторичночувствующих рецепторов между сенсорным нейроном и стимулом расположена рецептирующая клетка, в которой под влиянием раздражителя идут процессы трансформации энергии раздражителя в процесс возбуждения. Но в этой клетке не возникает импульсной активности. Рецепторные клетки синапсами соединены с сенсорными нейронами. Под влиянием потенциала рецептирирующей клетки выделяется медиатор, который возбуждает нервное окончание сенсорного нейрона и вызывает в нем появление локального ответа - постсинаптического потенциала. Он оказывает деполяризующее действие на отходящее нервное волокно, в котором возникает импульсная активность.
Следовательно, у вторичночувствующих рецепторов локальная деполяризация возникает дважды: в рецептирующей клетке и в сенсорном' нейроне. Поэтому принято называть градуальный электрический ответ рецептирующей клетки рецепторным потенциалом, а локальную деполяризацию сенсорного нейрона генераторным потенциалом, имея в виду, что он генерирует в отходящем от рецептора нервном волокне распространяющееся возбуждение. У первичночувствующих рецепторов рецепторный потенциал является и генераторным. Таким образом, рецепторный акт можно изобразить в виде следующей схемы.
Для первичночувствующих рецепторов:
I этап - специфическое взаимодействие раздражителя с мембраной рецептора;
II этап - возникновение рецепторного потенциала в месте взаимодействия раздражителя с рецептором в результате изменения проницаемости мембраны для ионов натрия (или кальция);
III этап - электротоническое распространение рецепторного потенциала к аксону сенсорного нейрона (пассивное распространение рецепторного потенциала вдоль нервного волокна называется электротоническим);
IV этап - генерация потенциала действия;
V этап - проведение потенциала действия по нервному волокну в ортодромном направлении.
Для вторичночувствующих рецепторов:
I-III этапы совпадают с такими же этапами первичночувствующих рецепторов, но протекают они в специализированной рецептирующей клетке и заканчиваются на ее пресинаптической мембране;
IV этап - выделение медиатора пресинаптическими структурами рецептирующей клетки;
V этап - возникновение генераторного потенциала на постсинаптической мембране нервного волокна;
VI этап - электротоническое распространение генераторного потенциала по нервному волокну;
VII этап - генерация потенциала действия электрогенными участками нервного волокна;
VIII этап - проведение потенциала действия по нервному волокну в ортодромном направлении.
Слуховые функции.
Анализ частоты звука (высоты тона). Звуковые колебания разной частоты вовлекают в колебательный процесс основную мембрану на всем ее протяжении неодинаково. Локализация амплитудного максимума бегущей волны на основной мембране зависит от частоты звука. Таким образом, в процесс возбуждения при действии звуков разной частоты вовлекаются разные рецепторные клетки спирального органа. В улитке сочетаются два типа кодирования, или механизма различения, высоты тонов: пространственный и временной. Пространственное кодирование основано на определенном расположении возбужденных рецепторов на основной мембране. Однако при действии низких и средних тонов, кроме пространственного, осуществляется и временное кодирование: информация передается по определенным волокнам слухового нерва в виде импульсов, частота следования которых повторяет частоту звуковых колебаний (см. рис. 14.14). О настройке отдельных нейронов на всех уровнях слуховой системы на определенную частоту звука свидетельствует наличие у каждого из них специфической частотно-пороговой характеристики — зависимости пороговой интенсивности звука, необходимой для возбуждения нейрона, от частоты звуковых колебаний. Для каждого нейрона существует оптимальная, или характеристическая, частота звука, на которую порог реакции нейрона минимален, а в обе стороны по диапазону частот от этого оптимума порог резко возрастает. При надпороговых звуках характеристическая частота дает и наибольшую частоту разрядов нейрона. Таким образом, каждый нейрон настроен на выделение из всей совокупности звуков лишь определенного, достаточно узкого участка частотного диапазона. Частотно-пороговые кривые разных клеток не совпадают, а в совокупности перекрывают весь частотный диапазон слышимых звуков, обеспечивая полноценное их восприятие.
Анализ интенсивности звука. Сила звука кодируется частотой импульсации и числом возбужденных нейронов. Увеличение числа возбужденных нейронов при действии все более громких звуков обусловлено тем, что нейроны слуховой системы отличаются друг от друга по порогам реакций. При слабом стимуле в реакцию вовлекается лишь небольшое число наиболее чувствительных нейронов, а при усилении звука в реакцию вовлекается все большее число дополнительных нейронов с более высокими порогами реакций. Кроме того, пороги возбуждения внутренних и наружных рецепторных клеток неодинаковы: возбуждение внутренних волос-ковых клеток возникает при большей силе звука, поэтому в зависимости от его интенсивности меняется соотношение числа возбужденных внутренних и наружных волосковых клеток.
Слуховые ощущения. Тональность (частота) звука. Человек воспринимает звуковые колебания с частотой 16—20 000 Гц. Этот диапазон соответствует 10—11 октавам. Верхняя граница частоты воспринимаемых звуков зависит от возраста человека: с годами она постепенно понижается и старики часто не слышат высоких тонов. Различение частоты звука характеризуется тем минимальным различием по частоте двух близких звуков, которое еще улавливается человеком. При низких и средних частотах человек способен заметить различия в 1—2 Гц. Встречаются люди с абсолютным слухом: они способны точно узнавать и обозначать любой звук даже при отсутствии звука сравнения.
Слуховая чувствительность. Минимальную силу звука, слышимого человеком в половине случаев его предъявления, называют абсолютным порогом слуховой чувствительности. Пороги слышимости зависят от частоты звука. В области частот 1000— 4000 Гц слух человека максимально чувствителен. В этих пределах слышен звук, имеющий ничтожную энергию. При звуках ниже 1000 и выше 4000 Гц чувствительность резко уменьшается: например, при 20 и при 20 000 Гц пороговая энергия звука в миллион раз выше (нижняя кривая AEFGD на рис. 14.16).
Усиление звука может вызвать неприятное ощущение давления и даже боль в ухе. Звуки такой силы характеризуют верхний предел слышимости (кривая ABCD на рис. 14.16) и ограничивают область нормального слухового восприятия.
[1] Раздражитель - любой материальный агент, внешний или внутренний, осознаваемый или неосознаваемый, выступающий как условие последующих состояний организма. Сигнальный раздражитель (он же индифферентный) - раздражитель, который прежде не вызывал соответствующей реакции, но при определенных условиях образования условного рефлекса, начинающий ее вызывать
[2] Временная связь - это совокупность нейрофизиологических, биохимических и ультраструктурных изменений мозга, возникающих в процессе сочетания условного и безусловного раздражителей и формирующих определенные взаимоотношения между различными мозговыми образованиями.
Дата добавления: 2015-11-04; просмотров: 7827;