Химические превращения, обеспечивающие сокращение и расслабление мышцы
Сокращение мышцы начинается с поступления в нервно-мышечный синапс двигательного импульса. Это вызывает освобождение в синапсе медиатора – ацетилхолина, под влиянием которого наступает возбуждение сарколеммы и открываются ионные каналы. Ионы натрия, концентрирующиеся в расслабленном волокне на поверхности сарколеммы и создающие положительный заряд, перемещаются на ее внутреннюю поверхность. При этом происходит нейтрализация отрицательного заряда, создаваемого на внутренней поверхности сарколеммы локализованными там ионами хлора. В результате деполяризации сарколеммы изменяется разность потенциалов между сарколеммой и внутренними участками мышечного волокна, следствием чего является образование потенциала действия. Возникающая при этом волна возбуждения быстро распространяется по трубчатым образованиям Т-системы внутрь мышечного волокна и достигает саркоплазматического ретикулума, открывая в нем Са2+-каналы и вызывая выход из его пузырьков в саркоплазму ионов кальция.
Повышение концентрации ионов Са2+ в зоне миофибрилл является пусковым механизмом для процесса их сокращения. Они активируют АТФ-азные центры в головках миозиновых нитей, которые катализируют гидролиз связанных с ними молекул АТФ на АДФ и неорганический фосфат, но продолжают удерживать оба продукта реакции (рис. 63, фазы 1→2). В результате происходит перераспределение энергии, заключенной в конечной фосфатной связи АТФ, что вызывает аллостерические изменения в головке миозина.
Одновременно ионы Са2+ устраняют блокаду активных центров на актиновых нитях со стороны тропонина, в следствие чего между головками миозиновых нитей и активными центрами актиновых нитей образуются поперечные соединения – «спайки». Возникает актомиозиновый комплекс (рис. 63, фазы 2→3). Это вызывает выброс АДФ и Н3РО4 из активного центра головок миозина и освобождение энергии. За счет освободившейся энергии происходят комформационные изменения головок миозиновых нитей, связанных с активными центрами актина: головки миозина изгибаются, принимая наклонное положение по отношению к оси миозиновых нитей. При этом они продвигают актиновые нити в направлении центра саркомера – совершают работу наподобие гребка весла (рис 63, фазы 3→4).
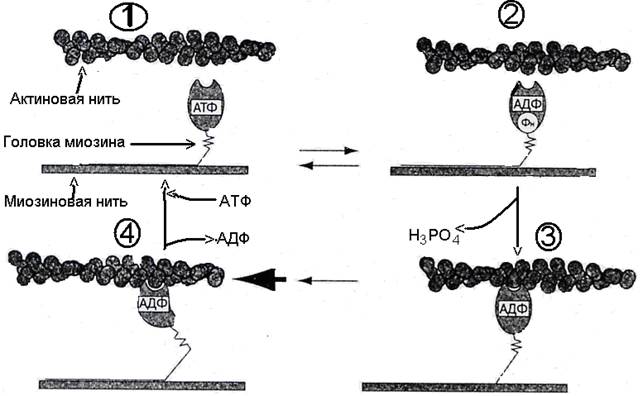
Рис. 63. Работа головок миозиновых нитей при мышечном сокращении
В таком положении головка миозина вновь связывает молекулу АТФ, «отпускает» активный центр актиновой нити и весь цикл повторяется. При каждом следующем цикле (гребке, шаге) головки миозина связываются с новыми активными центрами актина, за счет чего и происходит взаимное скольжение актиновых и миозиновых филаментов, т.е сокращение мышечного волокна.
Каждый «шаг» головок миозина обеспечивает взаимное смещение филаментов на 10 нм. Следовательно, полное сокращение мышцы обеспечивается огромным количеством повторных циклов. Частота «гребков» при быстром сокращении мышцы может достигать 5 раз в секунду.
По завершении сокращения (прекращении поступления двигательных импульсов) восстанавливается поляризация сарколеммы, концентрация ионов Са2+ в протоплазме снижается за счет активного обратного транспорта внутрь саркоплазматического ретикулума и его химического связывания, АТФ-азная активность миозина резко снижается, комплекс тропонин-тропомиозин занимает исходное положение на актине, блокируя место связывания с миозином. Результатом этого является расслабление мышцы.
Многие двигательные характеристики скелетных мышц хорошо согласуются с особенностями их молекулярного строения, а также структурными и химическими изменениями при сокращении. Так, напряжение, которое может развить мышца при сокращении, четко связано со степенью укорочения. Вначале эта зависимость прямая – чем больше укорочение, тем выше развиваемое усилие. Так продолжается пока длина мышцы не достигнет примерно 60% от исходной. Дальнейшее укорочение сопровождается снижением развиваемого усилия.
Причину этого явления легко понять, если рассмотреть происходящие в мышце процессы на молекулярном уровне. Развиваемое усилие напрямую зависит от количества «спаек» между головками миозина и актиновыми нитями, образующихся в каждый момент сокращения. С началом сокращения количество таких спаек непрерывно увеличивается, что и обуславливает увеличение развиваемого усилия. При значительной степени сокращения тонкие нити могут не только сходиться в зоне Н, но и перекрывать друг друга, препятствуя образованию спаек. При этом усилие, развиваемое мышечным волокном, уменьшается (рис.62).
Вопросы и задания для самоконтроля
1. Каково содержание воды, белков, липидов, углеводов и минеральных соединений в мышечной ткани?
2. Какие макроэргические соединения и другие источники энергии содержатся в мышечной ткани, какова их концентрация и локализация?
3. Как влияют систематические занятия физическими упражнениями на химический состав мышечной ткани?
4. Назовите важнейшие белки мышечной ткани. Каково их содержание, свойства, структурная организация и роль?
5. Назавите важнейшие структурные компоненты мышечной ткани и их роль в обеспечении мышечной работы.
6. Каково молекулярное строение сократительных нитей мышечного волокна – миофибрилл?
7. Какие существуют типы мышечных волокон? Чем оразличаются мышечные волокна разных типов?
8. Каков механизм мышечного сокращения?
9. Какие химические реакции обеспечивают мышечное сокращение?
10. Какие химические превращения происходят при расслаблении мышц?
11. Какова роль АТФ в двухфазной мышечной деятельности?
Дата добавления: 2015-10-29; просмотров: 2682;