Биосинтез ДНК
Удвоение ДНК у эукариот проходит в S-фазу клеточного цикла. Инициацию репликации регулируют специфические сигнальные белковые молекулы – факторы роста. Они связываются с рецепторами клеточных мембран, генерируя сигнал, который и побуждает клетку к началу репликации. Одними из первых активируются гены, кодирующие белки циклины. Циклинзависимые киназы, связывая циклин, переходят в активную форму и фосфорилируют специфические белки, которые регулируют синтез ферментов, обеспечивающих репликацию.
Синтез новых цепей ДНК может произойти только при расхождении родительских цепей. В точке начала репликаци (сайты инициации или ориджины) происходит локальное расхождение цепей ДНК и образуются две репликативные вилки, движущиеся в противоположных направлениях.
В образовании репликативной вилки принимает участие ряд белков и ферментов (Рис. 6.1.):
· семейство ДНК-топоизомераз обеспечивает устранение суперспирализации.
· ДНК-хеликазы,используя энергию АТФ, осуществляют разрыв водородных связей между полинуклеотидными цепями и расплетают двойную спираль ДНК.
В поддержании этого участка ДНК в раскрученном состоянии участвуют ДНК-связывающие белки (ДСБ). Они связываются с одноцепочечной ДНК по всей длине разделившихся цепей, предотвращая их комплементарное взаимодействие.
Репликация ДНК осуществляется ДНК-зависимыми ДНК-полимеразами. Субстратами и одновременно источниками энергии для синтеза служат дАТФ, дГТФ, дЦТФ и дТТФ. Ферменты проявляют каталитическую активность только в присутствии предварительно раскрученной матричной двухцепочечной ДНК. Синтез цепей происходит в направлении 5¢®3¢ растущей цепи. Матричная цепь всегда считывается в направлении 3¢®5¢, т. е. синтезируемая цепь всегда антипараллельна матричной цепи. В ходе репликации образуются 2 дочерние цепи, представляющие собой копии матричных цепей.
В синтезе эукариотических ДНК принимают участие 5 ДНК-полимераз. ДНК-полимераза g обеспечивает репликацию только митохондриальной ДНК. ДНК-полимеразы a, b, d, e участвуют в синтезе ДНК в ядре клеток.
Инициирует репликацию ДНК-полимераза a. Фермент обладает сродством к определенному сайту одноцепочечной ДНК. Присоединяясь к нему, ДНК-полимераза синтезирует небольшой фрагмент РНК – праймер, состоящий из 8-10 рибонуклеотидов, к которому присоединяет еще около 50 дезоксирибонуклеотидов. Таким образом, ДНК-полимераза a синтезирует олигонуклеотид, состоящий из короткой последовательности РНК и фрагмента цепи ДНК.
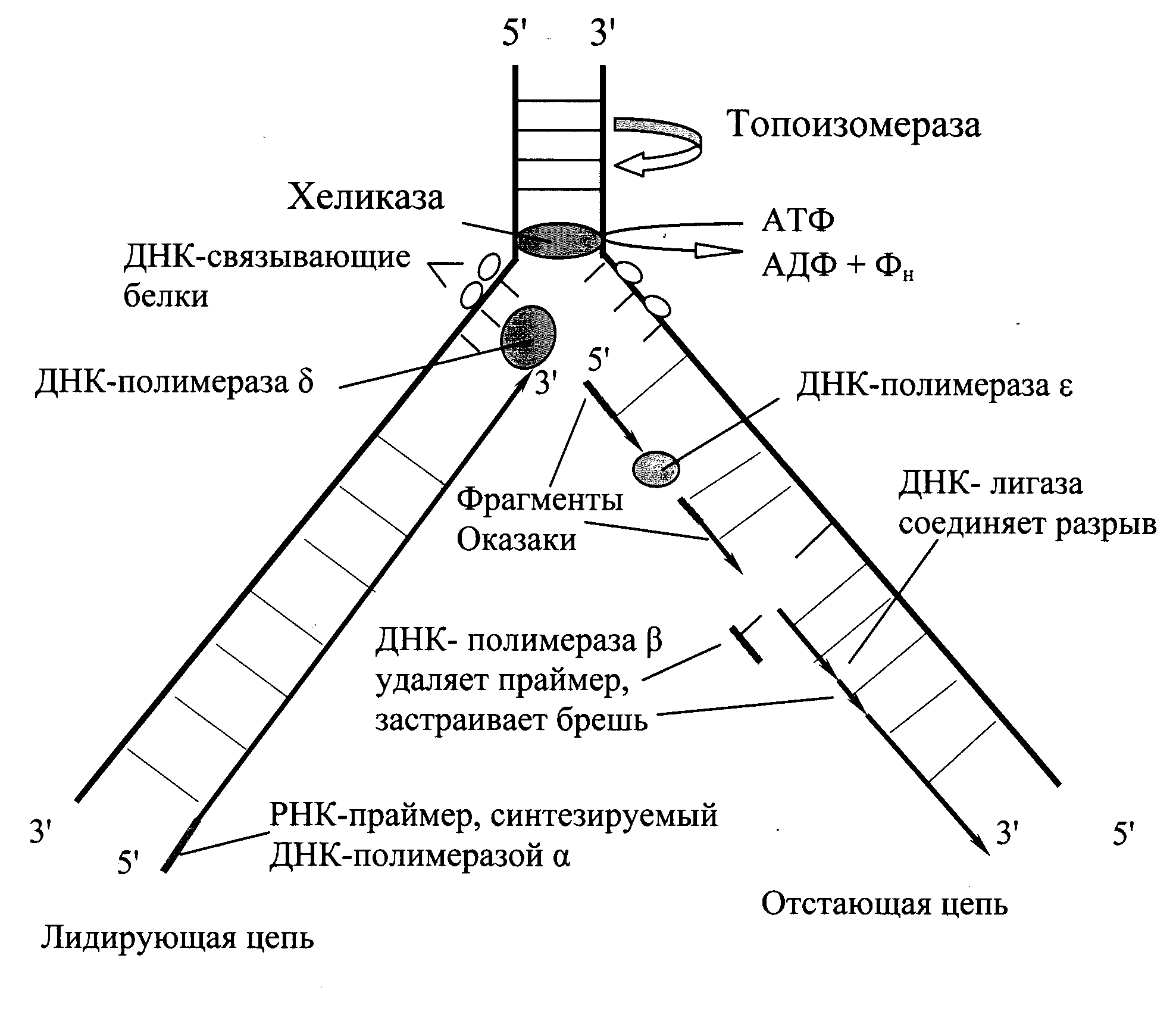
Рис. 6.1. Репликация ДНК
Олигонуклеотид, синтезированный ДНК-полимеразой a и образующий небольшой двухцепочечный фрагмент с матрицей, позволяет присоединиться ДНК-полимеразе d и продолжить синтез новой цепи в направлении 5¢®3¢ по ходу раскручивания репликативной вилки. Выбор ДНК-полимеразой очередного нуклеотида определяется матрицей: включение нуклеотида в синтезируемую цепь ДНК невозможно без предварительного связывания азотистого основания водородными связями с комплементарным нуклеотидом матричной цепи.
В каждой репликативной вилке идет одновременно синтез двух дочерних цепей. Направление синтеза цепи ДНК совпадает с направлением движения репликативной вилки лишь для одной из вновь синтезируемых цепей (лидирующая цепь). На второй матричной цепи синтез новой цепи осуществляется двумя ферментами: ДНК-полимеразой a и ДНК-полимеразой eв направлении 5¢®3¢, но противдвижения репликативной вилки. Поэтому вторая цепь синтезируется прерывисто, короткими фрагментами, которые по имени открывшего их исследователя называют «фрагменты Оказаки».Дочернюю цепь, синтез которой происходит фрагментами, а потому отстает, называют отстающей цепью.
Каждый фрагмент Оказаки содержит праймер. Праймеры удаляет ДНК-полимераза b,после чего присоединяет к ОН-группе на 3¢-конце предыдущего фрагмента дезоксирибонуклеотиды в количестве, равном вырезанному фрагменту и таким образом заполняет брешь, возникающую при удалении рибонуклеотидов.
ФерментДНК-лигазакатализирует образование фосфодиэфирной связи между 3¢-ОН-группой дезоксирибозы одного фрагмента и 5¢-фосфатом следующего. Реакция протекает с затратой энергии АТФ. Таким образом из множества фрагментов Оказаки образуется непрерывная цепь ДНК.
Терминация синтеза ДНК наступает вследствие исчерпания матрицы при встрече двух репликативных вилок.
После окончания репликации происходит метилирование вновь образованных цепей ДНК. Наличие СН3-групп необходимо для формирования структуры хромосом, а также для регуляции транскрипции генов.
На каждом конце хромосомы имеются неинформативные повторяющиеся последовательности нуклеотидов – теломеры.В соматических клетках с каждым актом репликации теломеры укорачиваются из-за невозможности достроить ДНК на месте 5¢-праймера. Это укорочение является важным фактором, определяющим продолжительность жизни клетки. Однако в эмбриональных и других быстро делящихся клетках потери концов хромосом недопустимы, так как укорочение хромосом будет происходить очень быстро. У эукариотических клетках имеется фермент теломераза, обеспечивающий восстановление недореплицированных 5¢-концов. В большинстве клеток теломераза неактивна, так как соматическая клетка имеет длину теломерной ДНК, достаточную для времени жизни клетки и её потомства. Небольшая активность теломеразы обнаруживается в клетках с высокой скоростью обновления, таких как лимфоциты, стволовые клетки костного мозга, клетки эпителия и т.д.
Дата добавления: 2015-10-05; просмотров: 1078;