Динамика популяций
8.5.1. Биотический потенциал
Любая популяция теоретически способна к неограниченному росту численности, если ее не лимитируют факторы внешней среды. В таком гипотетическом случае скорость роста популяции будет зависеть только от величины биотического потенциала, свойственного виду. Понятие биотического потенциала введено в экологию в 1928 г. Р.Чепменом. Этот показатель отражает теоретический максимум потомков от одной пары (или одной особи) за единицу времени, например за год или за весь жизненный цикл.
При расчетах его чаще всего выражают коэффициентом г и вычисляют как максимально возможный прирост популяции ΔN за отрезок времени Δt, отнесенный к одной особи, при начальной численности популяции N0:
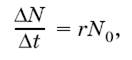
откуда
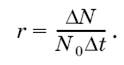
Величина биотического потенциала чрезвычайно различна у разных видов. Например, самка косули способна произвести за жизнь 10–15 козлят, трихина (Trichinella spiralis) – отложить 1,8 тыс. личинок, самка медоносной пчелы – 50 тыс. яиц, а луна-рыба – до 3 млрд икринок. Если бы все зародыши сохранялись, а все потомство выживало, численность любой популяции через определенные интервалы увеличивалась бы в геометрической прогрессии.
Кривая, отражающая на графике подобный рост популяции, быстро увеличивает крутизну и уходит в бесконечность (рис. 122). Такая кривая носит название экспоненциальной. На логарифмической шкале подобная зависимость численности популяции от времени будет представлена прямой, а биотический потенциал г отразится ее наклоном по отношению к горизонтальной оси, который тем круче, чем больше величина r.
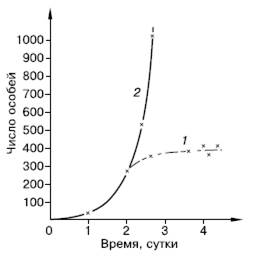
Рис. 122. Реальная (1) и теоретическая (2) кривая роста популяции парамеций
В природе биотический потенциал популяции никогда не реализуется полностью. Его величина обычно складывается как разность между рождаемостью и смертностью в популяциях: r = b – d, где b– число родившихся, а d – число погибших особей в популяции за один и тот же период времени.
Общие изменения численности популяции складываются за счет четырех явлений: рождаемости, смертности, вселения и выселения особей (иммиграция и эмиграция).
8.5.2. Рождаемость
Рождаемость – это число новых особей, появляющихся в популяции за единицу времени в расчете на определенное число ее членов.
Различают абсолютную и удельную рождаемость. Первая характеризуется общим числом родившихся особей. Например, если в популяции северных оленей, насчитывающей 16 тыс. голов, за год появилось 2 тыс. оленят, то это число и выражает абсолютную рождаемость. Удельную вычисляют как среднее изменение численности на особь за определенный интервал времени, и в данном случае она составит 0,125, т. е. один новорожденный на 8 членов популяции за год.
Величина рождаемости зависит от многих причин. Большое значение имеет доля особей, способных в данный период к размножению, что определяется соотношением полов и возрастных групп. Важна также частота последовательности генераций. Так, среди насекомых различают моновольтинные и поливольтинные виды. Первые дают одну, вторые – несколько генераций за год. Например, у тлей за сезон насчитывается до 15 партеногенетических поколений. По числу периодов размножения в течение жизни различают моноциклические и полициклические виды. Моноцикличность, или однократное размножение, свойственна обычно видам с короткой продолжительностью жизни в половозрелом состоянии (лососевые рыбы, поденки, майские жуки и многие другие насекомые). Полицикличность характеризуется повторным размножением особей и присуща большинству позвоночных животных и ряду беспозвоночных, например ракообразным.
У растений выделяют монокарпические и поликарпические виды, т. е. с однократным и многократным размножением в течение жизни. Для величины рождаемости имеет значение также соотношение периода размножения и общей продолжительности жизни. Этот период для самок дрозофил составляет около 65 %, для стадной саранчи шистоцерки – 15 %, а для поденок – всего от 0,5 до 1 % длительности их существования. Большую роль играет плодовитость особей. Однако размножаемость популяции, как правило, не бывает прямо пропорциональна плодовитости. Плодовитость сильно зависит от степени развития заботы о потомстве или обеспеченности яиц питательными материалами. Среди рыб наибольшее количество икринок выметывают виды с пелагической икрой – сельди, тресковые, камбаловые и др. Например, сахалинская сельдь продуцирует 38–46 тыс. мелких, в доли миллиметра, икринок. У лососевых, зарывающих икру в грунт, развивается меньшее число яиц, но более крупных размеров. Средняя плодовитость амурской горбуши 1300–1500 икринок диаметром 4–6 мм. Наиболее крупная икра у акул и химер, до 6–8 см. Яйца этих рыб имеют к тому же плотную защитную оболочку. Количество их очень невелико – несколько штук на одну самку.
У видов, оберегающих и выкармливающих свой молодняк, плодовитость резко понижена. Размер кладки у птиц разных видов варьирует уже не в тысячи раз, как у рыб, а в пределах от одного яйца (у некоторых хищников, пингвинов, чистиков и др.) до 20–25 (максимальное количество яиц у куриных, например, у серой куропатки).
Д. Лэ к установил на примере птиц, что у них отбор благоприятствует не максимальной плодовитости, а наиболее эффективной, т. е. тому количеству яиц, при котором потомство оказывается наиболее жизнеспособным. На выкармливание птенцов птицы тратят огромное количество энергии. Мелкие птицы приносят пищу в гнездо сотни раз в сутки: горихвостка – более 200, большая синица – около 400, а крапивник – до 600 раз. Если величина кладки больше обычного предела, птенцы оказываются недокормленными и их жизнеспособность снижена.
Большая плодовитость вырабатывается у видов также в условиях более высокой смертности, особенно при сильном давлении со стороны хищников. Отбор на плодовитость компенсирует высокую норму гибели в популяциях. Поэтому при высокой плодовитости рост численности популяции может быть весьма низким. В различных популяциях одного вида плодовитость обычно тем выше, чем более неблагоприятны занимаемые ими места обитания. Так, у многих млекопитающих – зайцев, мышей, полевок – число детенышей в помете на границах ареала больше, чем в его центре.
8.5.3. Смертность
Смертность в популяциях также зависит от многих причин: генетически запрограммированной длительности жизни особей, их генетической и физиологической полноценности, влияния неблагоприятных физических условий среды, воздействия хищников, паразитов, болезней и т. п. На разных стадиях жизненного цикла каждого поколения эти факторы действуют с разной силой. На практике, когда необходимо проанализировать ход смертности в популяциях, составляют таблицы выживания (см. далее), где для каждой возрастной группы указываются эмпирически полученные данные, характеризующие гибель особей данного возраста. На основе таких таблиц составляют кривые выживания, позволяющие прогнозировать в сходных условиях состояние очередных генераций.
Таблица 1
Таблица выживания куропатки (по Швердпфегеру, 1968)
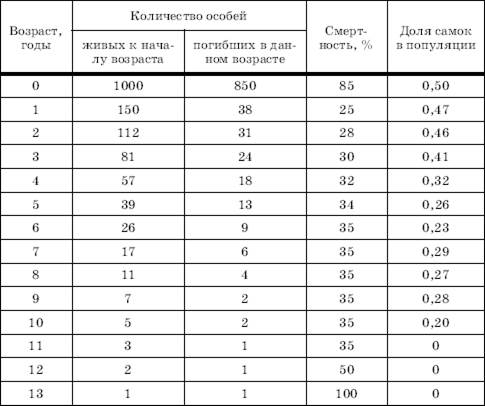
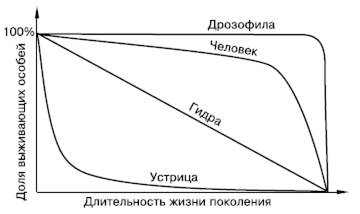
Рис. 123. Различные типы кривых выживаемости
Идеальный случай, если все особи одной генерации доживают до биологически предельного возраста, а затем в течение короткого срока отмирают. Кривая, описывающая такую зависимость численности поколений от времени, сначала идет параллельно горизонтальной оси, а затем круто загибается вниз (рис. 123). В природе подобный ход смертности не свойствен ни одному виду, но встречается приближение к нему, например, у таких насекомых с хорошо защищенными личинками, как жуки-златки, у крупных млекопитающих с высокой степенью выживаемости молодняка. К этому же типу близка кривая, описывающая смертность в современных популяциях человека. Средняя продолжительность жизни особи в таких популяциях высока и приближается к максимальной.
Наиболее часто встречающийся в природе вариант – повышенная гибель особей в ранний период жизни. Взрослые формы более защищены или выносливы. Кривая смертности в таких популяциях резко падает к горизонтальной оси в самом начале. Ее наклон отражает скорость убыли поколения. Так, у большинства рыб значительная часть популяции гибнет еще на стадии икры, очень велик также отсев мальков и лишь среди взрослых особей смертность становится ниже. У мышей и полевок, выкармливаемых матерью, гибель учащается после выхода молодых из гнезда, когда они приступают к самостоятельной жизни. Средняя продолжительность жизни особи намного меньше максимальной. Такая же картина характерна для многих воробьиных птиц (рис. 124). У людей также на протяжении почти всей истории была высокой детская смертность, которая резко снизилась с успешным развитием медицины. Это привело к изменению типа кривой выживаемости в поколениях и стремительному увеличению численности населения Земли, получившему название «демографического взрыва».
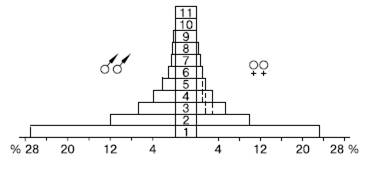
Рис. 124. Возрастная пирамида популяции зяблика Fringilla coelebs на Куршской косе: 1-11– возраст птиц, годы (из В. А. Паевского, 1985)
Сравнительно редко наблюдается третий вариант изменения выживаемости поколений – относительно равномерный отсев из-за случайных причин на протяжении всего жизненного цикла, без резко выраженных критических периодов повышенной смертности. На графике это выражается прямой линией между начальной и нулевой численностью генерации. Такой ход смертности отмечен, например, у пресноводных гидр, содержащихся в аквариуме.
8.5.4. Стратегии выживания популяций
Различия в биотическом потенциале видов зависят от их размеров, систематической принадлежности и других причин, но при прочих равных условиях связаны зависимостью со смертностью в популяциях. Эта закономерность, подмеченная еще Ч. Дарвином, была обоснована в трудах академика И. И. Шмальгаузена в 40-е годы прошлого столетия. Если вид подвергается в природе массовой неизбирательной элиминации, т. е. гибели от многочисленных врагов, избежать которых он бессилен, или подавляется другими экстремальными обстоятельствами, то единственным направлением отбора становится повышение размножаемости. В этом случае увеличивается вероятность случайного сохранения потомства и вид избегает вымирания. При неизбирательной элиминации различия между особями не имеют значения для их выживания, поскольку мощность воздействия губительных факторов слишком высока. При избирательной элиминации, когда смертность во многом определяется различиями между особями, отбор совершенствует разные формы морфофизиологических адаптаций, повышающих сопротивляемость вида влиянию неблагоприятных условий. Таким образом, высокий биотический потенциал – эволюционный ответ вида на пресс неблагоприятных для него воздействий среды, вызывающих высокую смертность.
В конце 60-х годов эта идея возродилась в концепции К– и г-отбора, выдвинутой американскими экологами Р. Макартуром и Э. Уилсоном. Они предложили различать две основные стратегии размножения организмов, обеспечивающие выживание в разных условиях, обозначив их через коэффициенты, входящие в уравнение роста популяций. При r-стратегии отбор идет на высокую плодовитость, оборачиваемость поколений, способность к быстрому расселению, что позволяет видам быстро восстанавливать численность после резкого ее снижения. При К-стратегии отбор совершенствует разные формы заботы о потомстве, что позволяет снизить плодовитость. Одновременно увеличивается продолжительность жизненных циклов и совершенствуются механизмы устойчивого поддержания численности в биоценозах. Естественно, что между крайними формами имеются все промежуточные варианты. Элементы К– и г-стратегий выживания прослеживаются во всех систематических группах организмов. Даже в пределах вида в популяциях, обитающих в разных условиях, усиливаются те или иные направления отбора.
8.5.5. Расселение
Выселение особей из популяции или пополнение ее пришельцами – закономерное явление, основанное на одной из важнейших биологических черт вида – его расселительной способности. В каждой популяции часть особей регулярно покидает ее, пополняя соседние или заселяя новые, еще не занятые видом территории. Этот процесс называют часто дисперсией популяции. Расселение приводит к занятию новых биотопов, расширению общего ареала вида, его успеху в борьбе за существование.
Расселительные функции выполняются в определенный период жизненного цикла: у насекомых в основном на стадии имаго, у тироглифоидных клещей – специальными дейтонимфами-гипопусами, у большинства птиц и млекопитающих – подрастающим молодняком. У растений рассеиваются или разносятся семена и споры, сидячие животные распространяются посредством плавающих личинок или специальных поколений при метагенезе.
Каждый вид характеризуется своим темпом дисперсии. По подсчетам, у зайцев-беляков регулярно покидает места рождения примерно 1 % молодняка, тогда как в популяциях большой синицы в среднем лишь треть молодых остается в той местности, где они вывелись из яиц.
Дисперсия обычно не направлена, расселение особей происходит в самых разных направлениях от мест отрождения.
Расселительная дисперсия служит средством связи между популяциями. Она повышается при увеличении плотности населения.
В период депрессии численности, наоборот, усиливается поток вселенцев в популяцию. У оседлых животных с хорошо выраженными территориальными инстинктами агрессивное поведение по отношению к пришельцам в период низкой численности популяции ослабевает, и вселенцы занимают свободные участки.
Ряд популяций, занимающих малопригодные места обитания, часто не в состоянии поддерживать свою численность за счет размножения и могут сохраняться преимущественно за счет иммиграции. Такие популяции В. Н. Беклемишев называл зависимыми.
Расселительные перемещения, по Н. П. Наумову, приводят к обмену особей между популяциями, увеличивают единство и общую устойчивость вида, так как те адаптации, которые возникли в местных условиях, но имеют общее значение, могут постепенно распространяться в пределах всего видового ареала. Проникновение расселяющихся особей на не занятые еще видом территории, заселение их и образование новых популяций называют инвазией.
8.5.6. Темпы роста популяции
Графики роста числа членов любой природной популяции в новых для нее условиях сильно отличаются от экспоненты. Кривая после подъема разной степени крутизны поворачивает параллельно горизонтальной оси, чем знаменует установление некоторой предельной численности популяции, которая затем поддерживается в течение более или менее длительного времени (рис. 122, 125). Такой ход кривой показывает, что в природе какие-то причины сдерживают чрезмерный рост популяции, не давая ей реализовать свой биотический потенциал, и ограничивают ее численность определенными пределами.
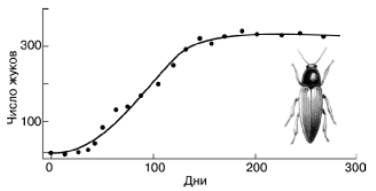
Рис. 125. Численность жуков в культуре, начатой с одной пары Rhizopertha dominica, в 10 г пшеницы (по Дж. Варли, 1978)
Пшеницу каждую неделю просеивали и дополняли до 10 г
Изменения темпов роста популяции могут быть различными.
В одном случае темп роста с самого начала высок и постоянен независимо от нарастающей плотности, что соответствует лавинообразному, по экспоненте, увеличению численности популяции. При достижении же определенной плотности населения темп роста падает сразу до нуля. Это означает, что популяция резко прекращает воспроизводство. В природе такой тип захвата среды встречается у видов, для которых жизненно важна скорость овладения ресурсами, например у нематод, клещей, некоторых насекомых, обитающих в быстро меняющих качество гниющих растительных остатках, кучках навоза копытных и т. п. Мелкие нематоды, населяющие конский и коровий навоз, могут осуществить свой жизненный цикл за несколько часов, тогда как близкие им виды, но обитающие в других, более постоянных условиях, развиваются от двух до трех недель. Скорость овладения средой имеет значение и для предупреждения конкуренции со стороны других претендентов. После массового размножения и быстрого потребления доступных ресурсов в популяциях подобных видов размножение прекращается и особи расселяются путем перелета на насекомых.
В другом случае изменения темпов роста популяций находятся в обратной зависимости от плотности. Разреженная популяция в благоприятных условиях быстро наращивает свою численность, но чем больше становится в ней особей, тем меньше доля очередного приплода, пока скорость воспроизводства не сравняется со смертностью (рис. 126). Тогда темп роста популяции снижается до нуля, а общая численность стабилизируется в соответствии с доступными для популяции ресурсами. Такой характер становления новых популяций свойствен в основном видам, у которых успех в размножении при низком уровне численности не лимитируется обязательностью группового образа жизни, необходимостью встречи полов и другими причинами (например, у мелких партеногенетических ракообразных, растений с апомиксисом и т. д.). Однако у большинства видов наиболее высокий темп роста популяций отмечается лишь при определенной оптимальной плотности. Если популяция сильно разрежена, это затрудняет встречу полов, защиту молодняка, проявление эффекта группы у животных, опыление у растений, поэтому население растет вначале очень медленно.
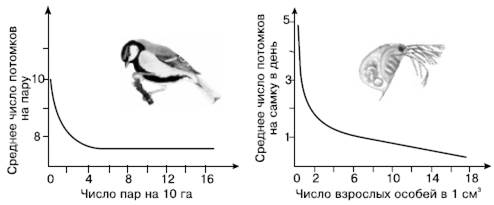
Рис. 126. Зависимость рождаемости от плотности в лабораторной популяции дафний и в дикой популяции большой синицы (по Ю. Одуму, 1975)
Как известно, появление потомства прежде всего зависит от числа производителей – особей, находящихся в генеративном возрастном состоянии. Вначале рост числа производителей сопровождается медленным ростом численности популяции, затем, в некотором интервале плотностей, зависимость выражена чрезвычайно резко, и даже небольшое увеличение доли производителей вызывает стремительный рост популяции, пока она не достигнет определенного уровня, который в последующем уже не меняется, сколько бы ни увеличивалась репродуктивная часть населения.
Такая зависимость была впервые предсказана французским математиком Верхюльстом в середине XIX века для населения людей, а позднее доказана англичанином Перлом (1925) для популяций животных в среде, где пищевые ресурсы имеют определенный предел восполнимости.
Установление определенного уровня плотности популяций после некоторого периода роста вовсе не означает, что в популяциях больше не происходит количественных изменений. Напротив, любое население всегда динамично и постоянно подвержено колебаниям численности, однако размах всех суточных, сезонных и годовых изменений популяций все же намного меньше теоретически возможного, соответствующего реализации всего биотического потенциала. Колебания численности совершаются с разным размахом вокруг некоторой средней величины, которая и соответствует горизонтальной части кривой на графике роста и стабилизации численности популяции.
Высокий потенциал размножения играет большую роль в выживании видов. Популяции, сведенные к низкому уровню численности, могут быстро восстановиться при благоприятной перемене условий. Некоторые виды только массовым размножением могут противостоять выеданию их различными потребителями или угрозе вытеснения конкурентами. Высокая размножаемость способствует быстрому освоению видом новых пространств.
Однако безграничное размножение таит в себе и большую опасность для любой популяции, так как может привести к быстрому подрыву ресурсов среды, нехватке пищи, убежищ, пространства и т. п., что неминуемо повлечет за собой общее ослабление популяции. Перенаселенность настолько неблагоприятна для любого вида, что в ходе эволюции у разных форм выработались в результате естественного отбора самые разнообразные механизмы, способствующие предотвращению избытка особей и поддержанию определенного уровня плотности популяций.
8.5.7. Динамика ценопопуляций растений
Все размерные и количественные характеристики растений в ценопопуляциях меняются в широком диапазоне.
Меняются такие показатели, как общая численность и плотность, фитомасса, площадь, занятая популяцией, проективное покрытие, возрастной спектр.
У большинства луговых растений колебания численности всходов как в течение сезона, так и по годам составляют от 1–2 до 100-1000 на м2. В степных ценозах в отдельные годы всходы могут не появиться совсем. Массовое их отмирание происходит, например, при наступлении засухи, поедании фитофагами, угнетении взрослыми растениями. Возрастные спектры ценопопуляций разных видов обладают разной степенью динамичности: 1) популяционные волны, (т. е. волны численности) перемещаются постепенно, при этом тип возрастного спектра не меняется и он остается полночленным. Это происходит при регулярном, но относительно небольшом пополнении молодыми особями; 2) популяционные волны перемещаются быстро, возрастной спектр может быть разорванным и неполночленным (рис. 127). Между этими двумя типами динамики различаются всевозможные переходы.
Большая лабильность всех популяционных показателей свойственна видам реактивным, эксплерентам способным очень быстро захватывать освободившиеся площади, но и освобождать их при вытеснении конкурентными видами.
Нередко наблюдается однонаправленное необратимое изменение ценопопуляции или ее отдельных локусов от зарождения до зрелости и старения. В конечном счете ценопопуляция или локус на данной площади исчезает. Такой тип динамики называется сукцессивным. Например, при интенсивной пастбищной нагрузке в ценопопуляциях мятлика однолетнего на залежах уменьшается плотность ценопопуляции, из нее выпадают молодые группы, происходит быстрое общее старение и ценопо-пуляция исчезает. В некоторых случаях сукцессивным типом динамики характеризуются отдельные локусы в пределах ценопопуляции, а сама она в целом сохраняет стабильность.
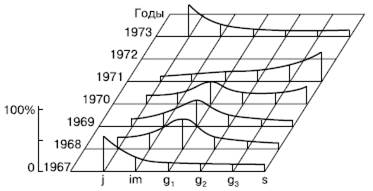
Рис. 127. Погодичные изменения популяционного потока у полевицы тонкой (по Е. И. Курченко, 1975)
В широколиственных лесах ценопопуляции луковичного растения гусиного лука желтого существуют в виде разобщенных локусов. Этот вид относится к группе эксплерентов, т. е. способен очень быстро захватывать освободившуюся территорию вследствие высокой скорости роста и большой энергии вегетативного размножения. Часто локус занят одним клоном, начало которому дает одна ювенильная особь. Перейдя в имматурное состояние, она начинает размножаться, образуя новые ювенильные особи. Значительная часть ювенильных растений переходит затем в покоящееся состояние, а нормально развивающиеся последовательно проходят все возрастные состояния вплоть до генеративного. В итоге все или почти все особи клона могут перейти в покоящееся состояние. Так завершается развитие локуса. Этот процесс длится 10–25 лет. Но в природе подобное происходит редко, так как даже незначительные нарушения почвы и лесной подстилки роющими животными приводят к пробуждению покоящихся луковичек. Развитие локуса опять начинается с ювенильного возрастного состояния растений, и процесс развития клонов приобретает циклический характер. Так как разные локусы развиваются асинхронно во времени и пространстве, вся ценопопуляция претерпевает флюктуационные изменения. Флюктуации – это обратимые, разнонаправленные изменения, когда чередуются периоды старения и омоложения ценопопуляции и поколения непрерывно сменяют друг друга. Таким образом популяция сохраняет за собой занятую площадь.
У некоторых луговых растений временное прекращение инспермации и одновременное взросление и старение особей может привести к тому, что возрастной спектр утратит молодую часть, станет разорванным, неполночленным (рис. 128). Когда возобновление восстановится, молодые особи при дальнейшем развитии популяции образуют новую популяционную волну, которая со временем сменит затухающую волну старой части ценопопуляций. Такие волнообразно-флюктуационные колебания численности и возрастной структуры прослежены, например, у луговика дернистого. Высокий динамизм наблюдается и в посевах луговых трав, особенно при действии различных антропогенных факторов, таких, как высокие дозы удобрений, полив, многократное отчуждение.
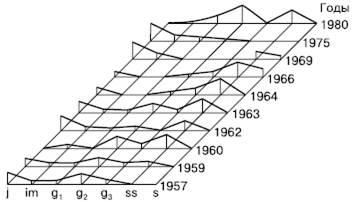
Рис. 128. Динамика ценопопуляций луговика дернистого на окских лугах (по Л. А. Жуковой, 1986)
8.5.8. Гомеостаз популяций
Поддержание определенной плотности получило название гомеостаза популяций. В основе способностей популяций к гомеостазу лежат изменения физиологических особенностей, роста, поведения каждой особи в ответ на увеличение или уменьшение числа членов популяции, к которой она принадлежит.
Механизмы популяционного гомеостаза зависят от экологической специфики вида, его подвижности, степени воздействия хищников и паразитов и др. У одних видов они могут проявляться в жесткой форме, приводя к гибели избытка особей, у других – в смягченной, например в понижении плодовитости на основе условных рефлексов.
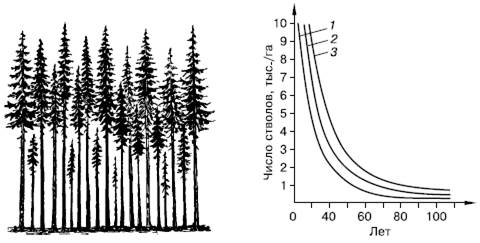
Рис. 129. Самоизреживание в древесных насаждениях (по Г. Ф. Морозову, 1928):
слева – господствующие и угнетенные деревья в ельнике; справа – ход изреживания стволов с возрастом у сосны (1), березы (2) и ели (3)
К жестким формам внутривидовой конкуренции следует отнести, например, явление самоизреживания у растений (рис. 129). При большой густоте всходов часть растений неминуемо погибает в результате угнетения физиологически более сильными соседями. Уменьшение числа растений происходит, даже если высеянные семена генетически однородны. В этом случае, по-видимому, имеют значение разница в размерах семян, во времени появления всходов, детали микроокружения. В одном из опытов с клевером Trifolium subterraneum через 84 дня после появления всходов на участке в 1 м2 из 1250 растений осталось 650, причем влияние вредителей было исключено.
У райграса многолетнего основной экологической единицей становится не особь, а побег. Обнаружено, что при разных нормах высева семян, от 6 до 180 кг/га, сначала густота побегов варьирует от 30 до 1070 на 100 см2, но затем во всех случаях становится равной примерно 500, т. е. в более редких посевах появляются новые побеги, а в более густых часть отмирает.
Регуляция плотности популяции у растений ввиду особенностей их роста происходит обычно не только путем изменения численности особей на единице площади, но и путем изменения вегетативной мощности каждого. В загущенных посевах растения менее облиственны, с меньшим количеством побегов. Общая их масса при увеличении плотности посевов сначала возрастает пропорционально количеству высеянных семян, а затем остается на постоянном уровне, тогда как средняя масса отдельных особей соответственно уменьшается. В данном случае стабилизируется не число особей в популяции, а общая листовая фотосинтезирующая поверхность растений.
У животных жесткие формы регуляции плотности популяций проявляются обычно лишь в тех случаях, когда запасы пищи, воды или других ресурсов резко ограничены, а животные либо не способны в данный период к поискам ресурсов на другой территории, либо эти поиски неэффективны. Например, в небольших пресноводных водоемах, где нет других видов рыб, популяции окуня могут поддерживать свое существование и регулировать плотность за счет питания взрослых собственной молодью. Мальки же растут за счет мелкого планктона, к питанию которым крупные окуни не приспособлены. Каннибализм – не частое явление в популяциях животных.
Особый интерес представляют некоторые, сравнительно редкие виды, у которых способность к умерщвлению конкурентов внутри популяции закрепляется эволюционно в их поведении и даже морфологии. Подобные примеры встречаются среди насекомых.
Для паразитоидных перепончатокрылых, откладывающих свои яйца в яйца или личинки других насекомых, запас пищи в хозяине весьма ограничен и дает возможность развиться чаще всего лишь одной особи паразита. У ряда видов личинки первого возраста имеют челюсти, ненужные при питании содержимым яйца, но приспособленные для схваток с конкурентами. Нападение личинок друг на друга обычно происходит до того, как они начинают кормиться. После уничтожения конкурента личинка линяет и ее острые челюсти сменяются мягкими и бугорковидными (рис. 130). У тех видов, самки которых способны распознать уже зараженных хозяев и не откладывать яйца в них повторно, личинкам не приходится конкурировать между собой, и они лишены острых челюстей.
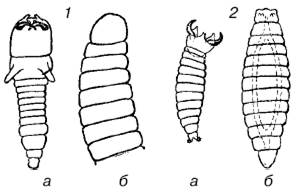
Рис. 130. Личинки паразитоидных перепончатокрылых:
1– Opius fletcheri; 2 – Galesus sylvestrii (a – первый возраст, б – второй возраст)
Групповые паразиты, откладывающие по нескольку яиц в одно насекомое, нередко обладают способностью соразмерять число яиц с величиной жертвы. Однако при недостатке хозяев возможно перезаражение обнаруженных паразитами, при этом обеспеченность потомства кормом снижается. В таких случаях наблюдается уменьшение размеров личинок. Вышедшие из них имаго откладывают вдвое-втрое меньше яиц при сокращенных сроках жизни. Например, у самых мелких наездников-браконид происходит полная редукция яичников.
Плотность популяции у паразитических перепончатокрылых может регулироваться также через изменения в соотношении полов, что влияет на численность следующего поколения. У многих видов сильно различается смертность самцов и самок при перенаселении. Например, у некоторых браконид доля самок составляет около 70 % при слабом заражении насекомых-хозяев и падает ниже 10 % – при сильном. Кроме того, соотношение полов регулируется поведением самок. Для многих видов наездников установлено, что в мелких хозяевах, представляющих малый запас корма, самки откладывают преимущественно неоплодотворенные яйца, из которых у перепончатокрылых развиваются самцы. При учащении контактов самок друг с другом или при восприятии следовых запахов, оставляемых другими самками, среди откладываемых яиц также увеличивается доля неоплодотворенных.
Среди механизмов, задерживающих рост популяций, у многих видов большую роль играют химические взаимодействия особей. Так, вода аквариума, в котором содержались дафнии, способна задерживать рост представителей того же вида и сохраняет эту способность в течение нескольких дней. Головастики выделяют в воду частицы белковой природы, которые задерживают рост других головастиков. Чем крупнее особь, тем сильнее она воздействует на более мелких, так как устойчивость к одной и той же концентрации ингибитора находится в прямой зависимости от размеров. Один крупный головастик Rana pipiens может задержать рост всех других в 75-литровом аквариуме. Поколение, вышедшее в близкие сроки из икры, отложенной в одном водоеме, вскоре разделяется на две размерные группы: более крупные, продолжающие расти головастики и мелкие, затормозившие свой рост из-за неблагоприятной для них концентрации метаболита. Экологическая выгода такого разделения популяции в том, что особи с наследственно более быстрым темпом роста, используя в полной мере кормовые ресурсы водоема, получают возможность быстро завершить метаморфоз и в популяцию вливается полноценное пополнение. Оставшиеся мелкие головастики, после того как первая партия покинет водоем и в нем снизится концентрация ингибитора, также имеют шансы увеличиться в размерах и достичь стадии метаморфоза, но значительно позже. Эту вторую часть пополнения можно рассматривать как своего рода резерв, который вливается в популяцию лишь при достаточно благоприятных условиях (если не пересохнет временный водоем, если в нем продолжится воспроизводство водорослей – основной пищи головастиков и т. п.).
Выделение в окружающую среду продуктов, задерживающих рост, обнаружено у многих растений и водных животных, особенно у рыб.
Другой механизм ограничения численности популяций – такие изменения физиологии и поведения при увеличении плотности, которые в конечном счете приводят к проявлению инстинктов массовой миграции. В результате происходит выселение большей части популяции за пределы территории, занимаемой в оседлый период. Особенно ярко это проявляется у насекомых, которым свойственна фазовость – резкое изменение морфологии и физиологии особей в зависимости от плотности популяции (рис. 131). У перелетной саранчи-шистоцерки в постоянных очагах ее обитания в Индии, Пакистане, Восточной Африке и Аравии при низкой численности личинки одиночной фазы имеют ярко-зеленую, а взрослые – серовато-зеленую или бурую окраску. В годы массового размножения, которое наступает при благоприятном сочетании погодных условий, саранча переходит в стадную фазу. Личинки приобретают ярко-желтую, с черными пятнами окраску, взрослая неполовозрелая шистоцерка интенсивно розово-лиловая, половозрелая – лимонно-желтая. Изменяется и морфология особей: удлиняются надкрылья, меняются формы переднеспинки, киля, пропорции конечностей и т. п. Переход из одной фазы в другую занимает около трех поколений. Процесс стимулируется зрительным восприятием особой своего вида и контактами при помощи усиков. Это вызывает в организме насекомых целую серию реакций, приводящих к гормональным перестройкам, в которых участвуют эндокринные железы.
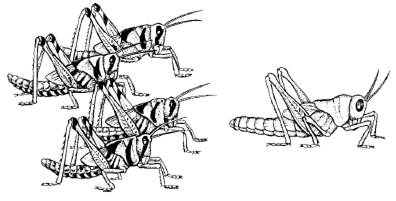
Рис. 131. Нимфы V возраста саранчи-шистоцерки (по Н. С. Щербиновскому, 1952): слева – стадная форма; справа – одиночная форма
Стадная фаза отличается повышенной возбудимостью и чрезвычайной прожорливостью. Плодовитость самок снижается, но они откладывают яйца с большим содержанием питательных веществ. Стадная саранча все время находится в состоянии миграционной активности. Личинки двигаются скоплениями – кулигами, а взрослые гигантскими стаями разлетаются на сотни и тысячи километров от мест постоянного обитания (рис. 132). Так, в конце прошлого столетия масса одной из стай шистоцерки, перелетевшей через Красное море, была определена не менее чем в 44 млн т.
На границах своего временного ареала перелетная саранча не может размножаться, и эти очаги вскоре затухают. Стаи или погибают, или, постепенно разреживаясь, начинают откочевку к зоне постоянных очагов. В изреженных популяциях вновь происходит переход к одиночной фазе, через 2–3 промежуточных поколения. Таким образом, разлет стай саранчи не обеспечивает формирования новых постоянных популяций, а практически служит лишь механизмом снятия перенаселенности в местах, благоприятных для размножения. При этом гибнет гигантское количество насекомых. Стаи, несущие на своем пути неисчислимые бедствия, сами оказываются обреченными.

Рис. 132. Нашествие перелетной саранчи
Явление фазовости обнаружено не только у стадных саранчовых, но и у других беспозвоночных. У тлей увеличение плотности населения вызывает появление крылатой фазы и разлет насекомых с образованием новых поселений. Обычно тли дают несколько поколений бескрылых самок, но в условиях постоянного перенаселения крылатые самки развиваются в каждом поколении. У ряда амеб химические изменения состава водной среды, вызванные переуплотнением популяции, стимулируют переход в подвижную жгутиковую стадию. В результате происходит быстрое рассредоточение особей в пространстве.
Территориальное поведение животных, выработавшееся в ходе эволюции как система инстинктов, – наиболее эффективный механизм сдерживания роста численности популяции на данной площади. Мечение и охрана участков, не допускающие размножения на них «чужих» особей, приводят к рациональному использованию территории. Избыточная часть популяции при этом не размножается или вынуждена выселяться за пределы занятого пространства. Это же относится и к выведенному потомству, среди которого лишь некоторая часть вследствие естественной смертности взрослых занимает освобождающиеся участки.
Выселения как ответная реакция на растущую плотность популяции свойственны многим видам птиц и млекопитающих. Кроме обычной расселительной дисперсии молодняка, для ряда видов с резкими колебаниями численности характерны массовые перемещения – нашествия. Они возникают нерегулярно, лишь в годы вспышек размножения, и не имеют постоянного направления. Такие нашествия описаны, например, у тундровых леммингов, белок Сибири и Северной Америки и др. При нашествиях часть особей остается на месте, а среди эмигрантов преобладают молодые.
Повышение плотности популяции может сопровождаться такими изменениями в физиологии особей, которые ведут к снижению рождаемости и увеличению смертности. У млекопитающих известно явление стресса, которое впервые было описано в 1936 г. физиологом Г. Селье для человека. В ответ на отрицательное воздействие каких-либо факторов в организме возникают реакции двух типов: 1) специфические, зависящие от природы повреждающего агента (например, усиление теплопродукции при действии холода), и 2) неспецифическая реакция напряжения как общее усилие организма приспособиться к изменившимся условиям. Эта общая реакция складывается из ряда физиологических и морфологических изменений, которые постепенно развертываются как единый процесс. Реакция напряжения, или стресс, возникает в ответ на любые отрицательные воздействия среды, в том числе и на отклонение плотности популяции от оптимума.
Большую роль в развитии стресса играют сигналы коры головного мозга, меняющие активность гипоталамуса – центрального звена вегетативной нервной системы. В свою очередь, деятельность гипоталамуса вызывает изменения в функционировании гипофизарно-надпочечниковой гормональной системы. В состоянии стресса у животных сильно увеличивается кора надпочечников и повышается концентрация кортикостероидных гормонов, выделяемых этим органом, а также происходит целый ряд других изменений в гормональном равновесии организма. У самок в популяции учащаются нарушения овуляции, резорбция эмбрионов, рано прекращается лактация, угасают инстинкты заботы о потомстве и т. п., уменьшается число выводков и количество в них молодых. В конечном счете все это приводит к торможению роста популяции. У мышевидных грызунов, содержащихся в клетках одинаковых размеров, четко проявляется обратно пропорциональная зависимость между численностью зверьков в клетке и массой их репродуктивных органов. В состоянии стресса у животных даже при достаточном снабжении кормом понижается устойчивость к вредным воздействиям среды, что увеличивает смертность.
От плотности населения зависит в первую очередь поведение животных. У многих видов в условиях скученности повышается уровень агрессивности, меняется реакция на особей противоположного пола, молодняк и т. д.
Стресс-реакция как механизм, регулирующий рождаемость, особенно отчетливо проявляется у животных с хорошо выраженной системой иерархического подчинения в группах.
Реакция напряжения свойственна подчиненным животным; у них также тормозится воспроизводительная функция. Доминирующие особи стресс-реакции не проявляют. В переуплотненных популяциях стресс распространяется на большую часть особей и, по-видимому, тормозит размножение.
Стрессовое состояние не вызывает необратимых изменений в половой системе, а приводит лишь к временному гормональному блокированию ее функции. После устранения перенаселенности способность к размножению может восстановиться в короткие сроки.
Закономерности стресса, вызываемого перенаселением, изучают в основном на лабораторных животных. Однако многочисленные факты, зарегистрированные в природных популяциях, позволяют предполагать, что в естественных условиях стресс играет немалую роль в регуляции численности и структуры популяций и поведения млекопитающих (рис. 133).
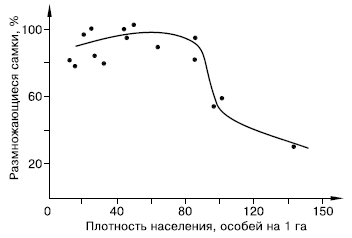
Рис. 133. Зависимость интенсивности размножения от плотности населения в популяции малого суслика (по М. Р. Магомедову, 1995)
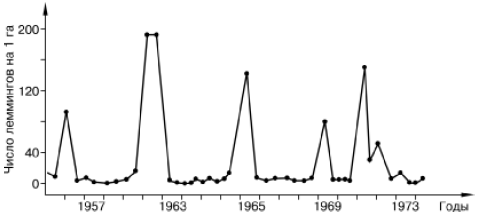
Рис. 134. Динамика численности сибирского лемминга на Аляске (по Bunnel et all., 1975)
Например, в динамике популяций ряда тундровых леммингов зарегистрированы правильные циклы трех-четырехлетней периодичности с амплитудой колебания до 600 крат (рис. 134). Фаза пика в таких колебаниях ограничена обычно одним сезоном, затем следуют резкий спад, фаза депрессии численности и последующего нарастания. При нарастании численности регистрируется увеличение плодовитости, повышение скорости созревания молодняка, усложнение возрастной структуры популяции, снижение смертности новорожденных. В период пика отмечается резкое снижение воспроизводства и одновременно увеличивается смертность во всех возрастных группах. В яичниках самок всех возрастов обнаруживаются серьезные деструктивные изменения, наблюдается массовая гибель фолликулов на ранних стадиях развития. Год-два после спада численности общая интенсивность размножения остается средней, а смертность высокой, а затем вновь все показатели размножаемости начинают расти. У зверьков, родившихся при низкой численности популяции, восстанавливается нормальное функционирование яичников.
На разных этапах этого цикла сильно меняется гормональное состояние зверьков, относящихся к разным генерациям. В пик численности отмечается избыточная активность адреналовой и щитовидной желез, что резко тормозит воспроизводительные функции организма. В ходе цикла у сменяющихся поколений меняется не только функционирование отдельных желез, но и всей эндокринной системы животных. Через несколько поколений, в период минимума численности, состояние эндокринной системы нормализуется и обеспечивает восстановление эффективности процесса размножения. Конкретные причины таких гормональных различий у разных поколений связаны с тем, что жизнеспособность и эндокринные особенности организма формируются в эмбриональный период и во многом определяются физиологическим состоянием родителей.
Таким образом, динамику численности леммингов можно представить как авторегулируемый процесс, в котором большую роль играют эндокринные механизмы.
Все рассмотренные выше примеры взаимодействия между членами популяции, от «жестких» форм – прямого уничтожения одной особью другой – до снижения воспроизводительных способностей как условного рефлекса на повышение частоты контактов, представляют собой разные формы ограничения роста популяций. Эти тормозящие механизмы включаются до полного истощения ресурсов среды в ответ на сигналы, свидетельствующие об угрозе перенаселения.
Степень развития механизмов популяционного гомеостаза находится также в тесной связи с тем, насколько влияют на популяцию другие виды: конкуренты, хищники, паразиты. Общая регуляция численности популяций в природных сообществах происходит в результате сложных межвидовых и внутривидовых взаимоотношений.
Дата добавления: 2015-09-23; просмотров: 1695;