Топологічні проблеми, пов'язані з реплікацією
Ще одним невід’ємним елементом системи реплікації є ДНК-топоізомерази. Рух реплікативної вилки з одночасним руйнуванням подвійної спіралі супроводжується прокруткою спіралі, що створює позитивні надспіральні витки – еластичні напруження попереду від вилки. Це стосується як еукаріотичних систем, де петлі ДНК зчеплені своїми кінцями з ядерним матриксом, так і прокаріотичних циркулярних хромосом: зустрічний рух двох реплікативних вилок (рис. 3) створює позитивні напруження на ділянці, яка ще не є реплікованою. Еластична напруга має час від часу зніматися, інакше врешті решт вона повністю зупинить рух реплісоми. Відповідно, процес реплікації потребує допоміжної дії топоізомераз, основна функція яких якраз і полягає в релаксації надспіралізованої ДНК: штучне вимикання активності топоізомераз повністю блокує реплікацію.
В еукаріотичній клітині завдання релаксації можуть виконувати як топоізомерази І, так і топоізомерази ІІ. Прокаріотична топоізомераза І (топоізомераза Іа) не підходить для виконання цієї ролі, оскільки здатна знімати тільки негативну надспіралізацію. Релаксація позитивної надспіралізації при реплікації у бактерій забезпечується гіразою – ферментом, що відноситься до класу топоізомераз ІІ. Гіраза вносить негативну надспіралізацію в циркулярну ДНК, що й компенсує позитивні напруження.
Ще одна топологічна проблема виникає при закінченні процесу реплікації в циркулярному репліконі. Коли дві реплікативні вилки зустрічаються, вони зупиняються за кілька витків подвійної спіралі одна від одної: еластичну напругу на маленькій ділянці зняти вже неможливо (рис. 17).
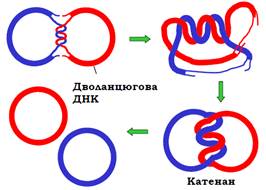
Рис. 17. Термінація реплікації у циркулярному репліконі.
Відбувається руйнування короткої подвійної спіралі, яка не піддалася реплікації, але це неможливо зробити без відповідного перекручення двох дочірніх кільцевих молекул ДНК десь у іншому місці. Одноланцюгові прогалини далі заповнюються ДНК-полімеразами. Але після цього дві остаточно синтезовані циркулярні молекули так і залишаються топологічно зчепленими одна з одною – утворюється так званий катенан (рис. 17). Таке саме перекручення двох дочірніх молекул ДНК реалізується при закінченні реплікації на двох сусідніх еукаріотичних репліконах (див. рис. 2).
Зрозуміло, що декатенація – розділення двох молекул – є неможливим без дволанцюгового розриву в одній із них. За цю операцію також відповідають топоізомерази класу ІІ (еукаріотична топоізомераза ІІ, яка зв’язана з ядерним матриксом під час реплікації, і гомологічна до неї топоізомераза ІV у прокаріотів), що роблять тимчасовий дволанцюговий розріз.
Дата добавления: 2015-09-11; просмотров: 1682;