Нуклеотиды
Нуклеотиды- фосфорные эфиры нуклеозидов.
Их химический состав: азотистое основание( А.О.) + пентоза + фосфорная кислота
Фосфорные эфиры образуются с участием гидроксильных групп пентоз . Места положения фосфорноэфирных групп принято обозначать, используя обозначение ( ' ) , например: 5' , 3 '
Предварительная краткая информация: нуклеотиды играют чрезвычайно важную роль в жизнедеятельности клетки.
Классификация нуклеотидов
Нуклеотиды, состоящие из одной молекулы А.О, пентозы, фосфорной кислоты, называются мононуклеотидами. Мононуклеотиды могут содержать одну молекулу фосфорной кислоты , две или три молекулы фосфорной кислоты, соединенных друг с другом.
Комбинация из двух мононуклеотидов называется динуклеотидом. Всоставе динуклеотида обычно присутствуют разные азотистые основания или одно другое циклическое соединение, например, витамин..
Особую роль в биохимических процессах играют циклические мононуклеотиды.
Номенклатура мононуклеотидов.
К названию нуклеозида добавляют в зависимости от количества фосфатных остатков, « монофосфат », « дифосфат », « трифосфат », с указанием их места положения в цикле пентозы- цифровое обозначение места со значком ( ' ) ,
Положение фосфатной группы в положении (5') является наиболее распространенным и типичным, поэтому его можно не указывать ( АМФ, ГТФ, УТФ, дАМФ и т.д.)
Остальные положения обозначаются обязательно ( 3'- АМФ, 2'- АМФ , 3'- дАМФ )
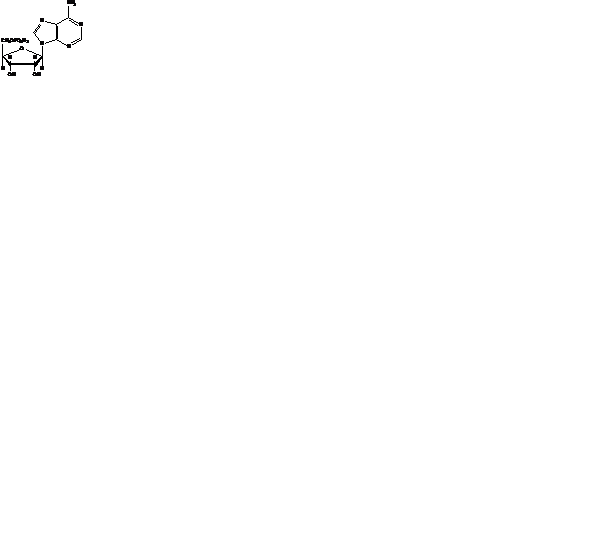
5'-аденозинмонофосфат
(5'- АМФ или АМФ )
Названия наиболее распространенных нуклеотидов
| нуклеозид | нуклеозидмонофосфат | нуклеозиддифосфат | нуклеозидтрифосфат |
| аденозин | 5'-Аденозинмонофосфат (5'- АМФ или АМФ ) 5' -адениловая кислота | 5'-Аденозиндифосфат ( 5'-АДФ или АДФ) | 5'-Аденозинтрифосфат ( 5'-АТФ или АТФ ) |
| аденозин | 3'-аденозинмонофосфат ( 3'-АМФ) 3' -адениловая кислота | не встречается in vivo | не встречается in vivo |
| гуанозин | 5'-гуанозинмонофосфат (5'- ГМФ или ГМФ ) | 5'-гуанозиндифосфат (5'- ГДФ или ГДФ ) | 5'-гуанозинтрифосфат (5'- ГТФ или ГТФ ) |
| гуанозин | 3'-гуанозинмонофосфат (3'- ГМФ) 3'-гуаниловая кислота | не встречается in vivo | не встречается in vivo |
| дезокси аденозин | 5'-дезоксиаденозин монофосфат (5'- дАМФ или дАМФ ) | 5'-дезоксиаденозин дифосфат (5'-дАДФили дАДФ) | 5'-дезоксиаденозин трифосфат (5'-дАТФили дАТФ) |
| уридин | 5'-уридинмонофосфат (5'- УМФ или УМФ) | 5'-уридиндифосфат (5'- УДФ или УДФ) | 5'-уридинтрифосфат (5'- УТФ или УТФ) |
| цитидин | 5'-цитидинмонофосфат (5'- ЦМФ или ЦМФ) | 5'-цитидиндифосфат (5'- ЦДФ или ЦДФ) | 5'-цитидинтрифосфат (5'- ЦТФ или ЦТФ) |
Нуклеотиды, образованные с участием рибозы, могут содержать остатки фосфорной кислоты в трех положениях ( 5', 3', 2' ), а с участием дезоксирибозы – только в двух положениях (5', 3' ) , в положении 2' гидроксигруппа отсутствует.. Это обстоятельство очень важно для структуры ДНК.
Отсутствие гидроксигруппы во втором положении имеет два важных последствия:
- уменьшается поляризация гликозидной связи в ДНК и она становится более устойчивой к гидролизу.
- 2-О-дезоксирибоза не может подвергаться ни эпимеризации, ни превращению в кетозу.
В клетке происходит последовательное превращение нуклеозидмонофосфата в дифосфат, а затем в трифосфат .
Для примера : АМФ ———> АДФ ———> АТФ
Биологическая роль нуклеотидов
Все нуклеозиддифосфаты и нуклеозидтрифосфаты относятся к высокоэнергетическим (макроэргическим ) соединениям.
Нуклеозидтрифосфаты участвуют в синтезе нуклеиновых кислот, обеспечивают активацию биоорганических соединений и биохимические процессы, которые проходят с затратой энергии. Аденозинтрифосфат ( АТФ) является наиболее распространенным в организме человека макроэргическим соединением. Содержание АТФ в скелетных мышцах млекопитающих до 4г/ кг, общее содержание около 125 г. У человека скорость обмена АТФ достигает 50 кг/ сутки. При гидролизе АТФ образуется аденозиндифосфат ( АДФ)

Макроэргические связи
В составе АТФ присутствуют разные типы химических связей:
- N -β- гликозидная
- сложноэфирная
- две ангидридные( в биологическом отношении макроэргические)
В условиях in vivo гидролиз макроэргической связи АТФ сопровождается выделением энергии( около 35 кДж/ моль), которая обеспечивает другие энергозависимые биохимические процессы.
АТФ + Н2О —фермент АТФгидролаза——> АДФ + Н3 РО4
В водных растворах АДФ и АТФнеустойчивы.При 00 САТФ стабильна в воде всего несколько часов, а при кипячении в течение 10 мин.
Под действием щелочи два концевых фосфата( ангидридные связи) гидролизуются легко, а последний( сложноэфирная связь) - трудно. При кислотном гидролизе N- гликозидная связь разрушается легко.
Впервые АТФ выделена из мышц в 1929 г. К. Ломаном. Химический синтез осуществил в 1948 г. А. Тодд.
Циклические нуклеотиды являются посредниками в передаче сигналов гормонов, изменяя в клетке активность ферментов.
Они образуются из нуклеозидтрифосфатов.
АТФ —фермент циклаза——> цАМФ + Н4 Р2 О7
После выполнения действия происходит гидролиз циклического нуклеотида. . Могут образоваться два соединения 5'- АМФ и 3' -АМФ, но в биологических условиях образуется только 5'-АМФ,

Циклический аденозинмонофосфат ( цАМФ)
11.5.Строение нуклеиновых кислот
Первичная структура РНК и ДНК – последовательное соединение нуклеотидов в полинуклеотидной цепи. Скелет полинуклеотидной цепи состоит из углеводных и фосфатных остатков , с углеводами посредством N- β – гликозидной связи соединены гетероциклические азотистые основания. С биологической точки зрения важнейшее значение имеют триплеты- блоки нуклеотидов из трех азотистых оснований, каждый из которых кодирует какую-либо аминокислоту или имеет определенную сигнальную функцию.
Структуру НК можно представить схематически :
5' 3' 5' 3' 5' 3'
фосфат —— пентоза —— фосфат —— пентоза —— фосфат —— пентоза -ОН
| | |
АО А О АО
В первичной структуре ДНК начало цепи определяют по пентозе, содержащей фосфат в положении 5'. Пентозы в полинуклеотидной цепи соединяются посредством фосфатных связей 3'→ 5'.На конце цепи в положении 3'- пентозы ОН- группа остается свободной.
Структура ДНК высшего порядка- двойная спираль
Научное описание вторичной структуры ДНК относится к величайшим открытиям человечества в ХХ веке. Биохимик Д. Уотсон и физик Ф. Крик в 1953 году предложили модель структуры ДНК и механизм процесса репликации. В 1962 г. им присуждена Нобелевская премия .
В популярном виде история описана в книге Джеймса Уотсона « Двойная спираль», М.: Мир, 1973 . Книга весьма интересно описывает историю совместной работы , с юмором и легкой иронией автора к такому знаменательному событию, счастливыми «виновниками» которого были два молодых ученых . С момента открытия структуры ДНК человечество получило инструмент к развитию нового направления- биотехнологиям, синтезу белков путем рекомбинации генов ( гормоны в медицинской промышленности получают инсулин, эритропоэтин и многие другие ).
Открытию структуры ДНК способствовали исследования Э.Чаргаффа в отношении химического состава ДНК. Он обнаружил:
- количество пиримидиновых оснований равно количеству пуриновых
- количество тимина равно количеству аденина, а количество цитозина количеству
гуанина.
А = Т Г = Ц
А + Г = Т + Ц
А + Ц = Т + Г
Эти отношения получили название правила Чаргаффа.
Молекула ДНК представляет собой две перекрученные спирали. Скелет каждой спирали- цепочка из чередующихся остатков дезоксирибозы и фосфорной кислоты. Спирали ориентированы таким образом., что образуют два неодинаковых спиральных желобка, которые идут параллельно главной оси. Эти желобки заполнены белками гистонами. Азотистые основания располагаются внутри спирали, почти перпендикулярно основной оси и образуют между цепочками комплементарные пары А…Т и Г…Ц.
Суммарная длина молекул ДНК в каждой клетке достигает 3 см. Диаметр клетки в среднем 10–5 м , диаметр ДНК всего 2 •10 –9 м.
Основные параметры двойной спирали:
* диаметр 1,8 – 2нм,
* на одном витке 10 нуклеотидов
* высота шага витка ~ 3,4 нм
* расстояние между двумя нуклеотидами 0,34 нм .
Основания располагаются перпендикулярно оси цепи.
* направления полинуклеотидных цепей антипараллельное
* связь между фуранозными циклами дезоксирибозы посредством
фосфорной кислоты осуществляется из положения 3` к положению 5` в
каждой из цепей.
* Начало цепи – фосфорилирована гидроксильная группа пентозы в положении
- 5`, конец цепи – свободная гидроксильная группа пентозы в положении 3`.
* В составе ДНК и РНК нуклеозидные фрагменты находятся в анти- конформации пиримидиновый цикл пурина находится справа от гликозидной связи. Только такое положение позволяет образовать комплементарную пару ( см. формулы нуклеотидов)
* Между азотистыми основаниями возникают три вида взаимодействий:
1. “Поперечное”, участвуют комплементарные пары двух цепей. Возникает «циклический» перенос электронов между двумя азотистыми основаниями (Т – А, У – Ц), образуется дополнительная p - электронная система, которая обеспечивает дополнительное взаимодействие и защищает азотистые основания от нежелательных химических воздействий. Между аденином и тимином устанавливается две водородные связи, а между гуанином и цитозином – три водородные связи.
2. « Вертикальное» (stacking) , за счет укладывания в “стопки” , участвуют азотистые основания одной цепи. «Стэкинг- взаимодействие» имеет даже большее значение в стабилизации структуры, чем взаимодействие в комплементарных парах
3. Взаимодействие с водой играет существенную роль в поддержании пространственного строения двойной спирали, которая принимает максимально компактную структуру для уменьшения поверхности контакта с водой и направляет гидрофобные гетероциклические основания вовнутрь спирали.
Структура и состав нуклеопротеидных комплексов
В связывании нуклеиновой кислоты с белком принимают участие несколько видов взаимодействия:
- электростатическое
- водородные связи
- гидрофобное
По результатам рентгеноструктурного анализа путем компьютерного моделирования построены реальные трехмерные модели ДНК, рибосом, информосом и нуклеиновых кислот вирусов.
Гистоновые белки ДНК обладают выраженными основными свойствами и отличаются высокой степенью эволюционной консервативности. По соотношению двух основных аминокислот лизин/ аргинин их подразделяют на 5 классов : Н1, Н2А, Н 2В, Н3, Н4
Дата добавления: 2015-08-11; просмотров: 2027;