Эксцизионная репарация
Эксцизионная репарация включает более сложные механизмы репарации, при которых “неправильные” участки ДНК вырезаются с помощью специальных ферментов. Рассмотрим некоторые типы эксцизионной репарации.
Удаление неправильного азотистого основания с образованием АР-сайтаосуществляется фрагментом ДНК-N-гликозилазой, которая разрушает гликозидную связь между азотистым основанием и дезоксирибозой, в результате чего вырезается только азотистое основание с сохранением сахарофосфатного остова – образуется АР-сайт (от слова апуринизация). Для дальнейшей репарации включаются дополнительные механизмы:
У эукариотобнаружен фермент инсертаза, который напрямую застраивает АР-сайт недостающим азотистым основанием, комплементарным соседней цепи.
У всех организмов может включаться сложный механизм эксцизии с помощью АР-эндонуклеаз, которые с 3\ или 5\-конца АР-сайта гидролизуют цепь ДНК. ДНК-полимераза І с помощью 3\ - или 5\ -экзонуклеазной активности убирает несколько нуклеотидов, включая АР-сайт, а с помощью полимеризующей активности застраивает образовавшуюся брешь. Лигаза сшивает концы фрагментов. Таким образом в ексцизионной репарации участвуют в основном ферменты репликации.
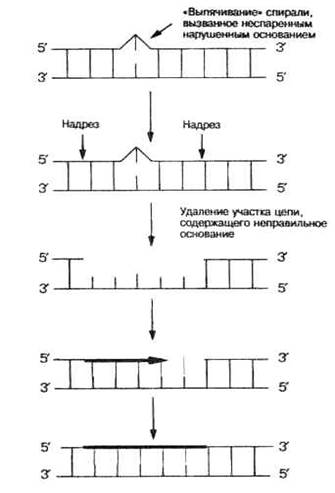
Рис.4.3. Механизм эксцизионной репарации
(аналогично происходит и темновая репарация)
Темновая репарацияявляется частным случаем эксцизионной репарации; однако она осуществляется специальными ферментами, которые кодируются генами uvr А, В, С, D. Эта репарация включается в темноте, когда не работает фотолиаза, или же когда в ДНК очень много УФЛ-повреждений. Эндонуклеаза uvr АВС разрывает фосфодиэфирные связи с 5\ и 3\ концов от поврежденного участка (иногда на значительном удалении), хеликаза ІІ при этом разводит цепи молекулы ДНК, а фермент uvr D выщепляет поврежденный фрагмент. ДНК-полимераза І застраивает образованную брешь, используя в качестве матрицы неповрежденную цепь, а лигаза сшивает фрагменты.
- Индуцированная SOS-репарация
Индуцированная SOS-репарация – один из наиболее сложных механизмов репарации. Обнаружена у вирусов, прокариот и эукариот. Ее суть сводится к индуцированному включению отдельных генов (у эукариот) или группы генов в опероне (у прокариот). В качестве индуктора выступает поврежденная ДНК (тиминовые димеры, одноцепочные ДНК, короткие фрагменты ДНК). SOS-система включается, когда в клетке много повреждений, а другие системы репарации не справляются.
Рассмотрим механизм SOS-репарации, обнаруженный у E. coli. Ферменты SOS-репарации кодируются 17 генами, собранными в один большой оперон, который работает по типу индуцибельного оперона с аутогенным контролем.
Этот оперон включается не сразу, а постепенно, как реостат, по мере нарастания количества поврежденной ДНК (индуктора). При отсутствии мутаций, транскрипция этого оперона блокируется белком-регулятором LexA, сидящим на операторе. Появляющиеся в результате повреждений фрагменты ДНК (индукторы) связываются с другим белком-регулятором RecA и этот комплекс (RecA + Индуктор) воздействует на LexA, вызывая его протеолиз. LexA отпадает от оператора и начинается транскрипция генов uvr АВСД с включением темновой репарации. Если для исправления мутаций этого оказывается достаточно, то экспрессированный далее LexA блокирует оператор. Если же поврежденная ДНК (индуктор) продолжает прибывать, то экспрессированный следом RecA активируется индуктором и снова убирает LexA с оператора. В результате этого транскрипция продолжается в области других генов и в том числе umuС и umuД, продукты которых ингибируют экзонуклеазную активность Е-субъединицы ДНК-полимеразы ІІІ. А именно эта субъединица ответственна за коррекцию синтеза. Таким образом, во избежание полной деградации ДНК SOS-система допускает даже неправильное встраивание нуклеотидов.
Дата добавления: 2015-08-08; просмотров: 2309;