Кровеносная система. Функции и свойства крови
Главным отличием кровеносной системы рыб от других позвоночных является наличие одного круга кровообращения и двухкамерного сердца, наполненного венозной кровью (за исключением двоякодышащих и кистёперых).
Сердце состоит из одного желудочка и одного предсердия и помещается в околосердечной сумке, сразу за головой, позади последних жаберных дуг, т. е. по сравнению с другими позвоночными сдвинуто вперед. Перед предсердием имеется венозная пазуха, или венозный синус, со спадающими стенками; через эту пазуху кровь поступает в предсердие, а из него – в желудочек.
Расширенный начальный участок брюшной аорты у низших рыб (акулы, скаты, осетровые, двоякодышащие) образует сокращающийся артериальный конус, а у высших рыб – луковицу аорты, стенки которой сокращаться не могут. Обратному току крови препятствуют клапаны.
Схема кровообращения в самом общем виде представлена следующим образом. Венозная кровь, заполняющая сердце, при сокращениях сильного мускульного желудочка через артериальную луковицу по брюшной аорте направляется вперед и поднимается в жабры по приносящим жаберным артериям. У костистых рыб их четыре с каждой стороны головы – по числу жаберных дуг. В жаберных лепестках кровь проходит через капилляры и, окисленная, обогащенная кислородом, направляется по выносящим сосудам (их также четыре пары) в корни спинной аорты, которые затем сливаются в спинную аорту, идущую вдоль тела назад, под позвоночником. Соединение корней аорты спереди образует характерный для костистых рыб головной круг. Вперед от корней аорты ответвляются сонные артерии.
От спинной аорты идут артерии к внутренним органам и мускулатуре. В хвостовом отделе аорта переходит в хвостовую артерию. Во всех органах и тканях артерии распадаются на капилляры. Собирающие венозную кровь венозные капилляры впадают в вены, несущие кровь к сердцу. Хвостовая вена, начинающаяся в хвостовом отделе, войдя в полость тела, разделяется на воротные вены почек. В почках разветвления воротных вен образуют воротную систему, а выйдя из них, сливаются в парные задние кардинальные вены. В результате слияния вен задних кардинальных с передними кардинальными (яремными), собирающими кровь из головы, и подключичными, приносящими кровь из грудных плавников, образуется два Кювьерова протока, по которым кровь попадает в венозный синус. Кровь из пищеварительного тракта (желудка, кишечника) и селезенки, идущая по нескольким венам, собирается в воротную вену печени, разветвления которой в печени образуют воротную систему. Собирающая кровь из печени печеночная вена впадает прямо в венозный синус (рис. 21). В спинной аорте радужной форели обнаружена эластичная связка, выполняющая роль нагнетающего насоса, который автоматически увеличивает циркуляцию крови во время плавания, особенно в мускулатуре тела. Производительность этого ‛дополнительного сердца“ зависит от частоты движений хвостового плавника.
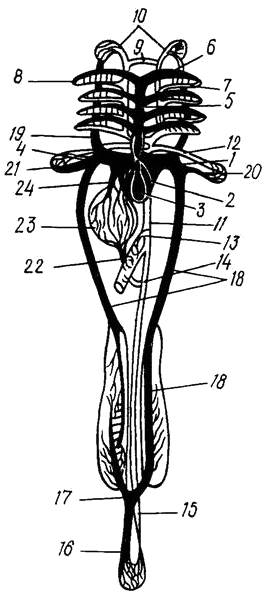
Рис. 21. Схема кровеносной системы костистой рыбы (по Наумову, 1980):
1 – венозная пазуха, 2 – предсердие, 3 – желудочек, 4 - луковица аорты, 5 – брюшная аорта,
6 – приносящие жаберные артерии, 7 – выносящие жаберные артерии, 8 – корни спинной аорты,
9 – передняя перемычка, соединяющая корни аорты, 10 – сонная артерия, 11– спинная аорта,
12 -подключичная артерия, 13 – кишечная артерия, 14 – брыжеечная артерия, 15– хвостовая артерия,
16 – хвостовая вена, 17– воротные вены почек, 18 – задняя кардинальная вена, 19 – передняя кардинальная вена, 20 – подключичная вена, 21 – Кювьеров проток, 22 – воротная вена печени,
23 – печень, 24 – печеночная вена; черным показаны сосуды с венозной кровью,
белым – с артериальной
У двоякодышащих рыб появляется неполная перегородка предсердия. Это сопровождается и возникновением ‛лёгочного“ круга кровообращения, проходящего через плавательный пузырь, превращенный в легкое.
Сердце рыб относительно очень мало и слабо, гораздо меньше и слабее, чем у наземных позвоночных. Масса его обычно не превышает 0,33–2,5%, в среднем 1 % массы тела, тогда как у млекопитающих оно достигает 4,6%, а у птиц даже 10–16%.
Кровяное давление (Па) у рыб низкое – 2133,1 (скат), 11198,8 (щука), 15998,4 (лосось), тогда как в сонной артерии лошади – 20664,6.
Невелика и частота сокращений сердца – 18–30 ударов в минуту, причем она сильно зависит от температуры: при низких температурах у рыб, зимующих на ямах, она уменьшается до 1–2 ;у рыб, переносящих вмерзание в лед, пульсация сердца на этот период прекращается.
Количество крови у рыб относительно меньше, чем у всех остальных позвоночных животных (1,1 – 7,3% от массы тела, в том числе у карпа 2,0–4,7%, сома – до 5, щуки – 2, кеты – 1,6, тогда как у млекопитающих – 6,8% в среднем). Это связано с горизонтальным положением тела (нет необходимости проталкивать кровь вверх) и меньшими энергетическими тратами в связи с жизнью в водной среде. Вода является гипогравитационной средой, т. е. сила земного притяжения здесь почти не сказывается.
Морфологическая и биохимическая характеристика крови различна у разных видов в связи с систематическим положением, особенностями среды обитания и образа жизни. Внутри одного вида эти показатели колеблются в зависимости от сезона года, условий содержания, возраста, пола, состояния особей.
Количество эритроцитов в крови рыб меньше, чем у высших позвоночных, а лейкоцитов, как правило, больше. Это связано, с одной стороны, с пониженным обменом рыб, а с другой – с необходимостью усилить защитные функции крови, так как окружающая среда изобилует болезнетворными организмами. По средним данным, в 1 мм3крови количество эритроцитов составляет (млн. ): у приматов –9,27; копытных– 11,36; китообразных – 5,43; птиц – 1,61–3,02; костистых рыб– 1,71 (пресноводные), 2,26 (морские), 1,49 (проходные).
Количество эритроцитов у рыб колеблется в широких пределах, прежде всего в зависимости от подвижности рыб: у карпа – 0,84–1,89 млн. /мм3 крови, щуки – 2,08, пеламиды–4,12 млн. /мм3. Количество лейкоцитов составляет у карпа 20–80, у ерша – 178 тыс. /мм3. Клетки крови рыб отличаются большим разнообразием, чем у какой-либо другой группы позвоночных. У большинства видов рыб в крови имеются и зернистые (нейтрофилы, эозинофилы) и незернистые (лимфоциты, моноциты) формы лейкоцитов. Среди лейкоцитов преобладают лимфоциты, на долю которых приходится 80–95%, моноциты составляют 0,5–11%; среди зернистых форм преобладают нейтрофилы–13–31%; эозинофилы встречаются редко (у карповых, амурских растительноядных, некоторых окуневых).
Соотношение разных форм лейкоцитов в крови карпа зависит от возраста и условий выращивания.
Общее количество лейкоцитов в крови рыб сильно изменяется в течение года, у карпа оно повышается летом и понижается зимой при голодании в связи со снижением интенсивности обмена.
Кровь окрашена гемоглобином в красный цвет, но есть рыбы и с бесцветной кровью. Так, у представителей семейства Chaenichthyidae (из подотряда нототениевых), обитающих в антарктических моряхв условиях низкой температуры (<2°С), в воде, богатой кислородом, эритроцитов и гемоглобина в крови нет. Дыхание у них происходит через кожу, в которой очень много капилляров (протяженность капилляров на 1 мм2 поверхности тела достигает 45 мм). Кроме того, у них ускорена циркуляция крови в жабрах.
Количество гемоглобина в организме рыб значительно меньше, чему наземных позвоночных: на 1 кг массы тела у них приходится 0,5–4 г, тогда как у млекопитающих этот показатель возрастает до 5–25 г. У быстра передвигающихся рыб обеспеченность гемоглобином выше, чем у малоподвижных (у проходного осетра 4 г/кг, у налима 0,5 г/кг). Количество гемоглобина в крови рыб колеблется в зависимости от сезона (у карпа повышается зимой и понижается летом), гидрохимического режима водоема (в воде с кислым значением рН, равным 5,2, количество гемоглобина в крови возрастает), условий питания (карпы, выращенные на естественной пище и дополнительных кормах, имеют разную обеспеченность гемоглобином). Ускорение темпа роста рыб коррелирует с повышенной обеспеченностью их организма гемоглобином.
Способность гемоглобина крови извлекать кислород из воды у разных рыб неодинакова. У быстро плавающих рыб – макрели, трески, форели – гемоглобина в крови много, и они очень требовательны к содержанию кислорода в окружающей воде. У многих морских придонных рыб, а также угря, карпа, карасей и некоторых других, наоборот, гемоглобина в крови мало, но он может связывать кислород из среды даже с незначительным количеством кислорода.
Например, судаку для насыщения крови кислородом (при 16°С) необходимо содержание в воде 2,1–2,3 О2 мг/л; при наличии в воде 0,56–0,6 О2 мг/л кровь начинает его отдавать, дыхание оказывается невозможным и рыба гибнет. Лещу при этой же температуре для полного насыщения гемоглобина крови кислородом достаточно присутствия в литре воды 1,0–1,06 мг кислорода.
Чувствительность рыб к изменениям температуры воды также связана со свойствами гемоглобина: при повышении температуры воды потребность организма в кислороде увеличивается, но способность гемоглобина связывать его падает.
Угнетает способность гемоглобина связывать кислород и углекислота: для того чтобы насыщенность кислородом крови угря достигла 50% при содержании в воде 1% СО2, необходимо давление кислорода в 666,6 Па, а в отсутствии СО2 для этого достаточно давления кислорода почти вдвое меньшего – 266,6– 399,9 Па.
Группы крови у рыб впервые были определены на байкальском омуле и хариусе в 30-х годах. К настоящему времени установлено, что групповая антигенная дифференцировка эритроцитов широко распространена; выявлено 14 систем групп крови, включающих более 40 эритроцитарных антигенов. При помощи иммуносерологических методов изучается изменчивость на разных уровнях; выявлены различия между видами и подвидами и даже между внутривидовыми группировками у лососевых (при изучении родства форелей), осетровых (при сравнении локальных стад) и других рыб.
Кровь, будучи внутренней средой организма, содержит в плазме белки, углеводы (гликоген, глюкоза и др.) и другие вещества, играющие большую роль в энергетическом и пластическом обмене, в создании защитных свойств. Уровень этих веществ в крови зависит от биологических особенностей рыб и абиотических факторов, а подвижность состава крови позволяет использовать ее показатели для оценки физиологического состояния.
Костного мозга, являющегося основным органом образования форменных элементов крови у высших позвоночных, и лимфатических желез (узлов) у рыб нет.
Кроветворение у рыб по сравнению с высшими позвоночными отличается рядом особенностей:
1. Образование клеток крови происходит во многих органах. Очагами кроветворения у рыб являются: жаберный аппарат (эндотелий сосудов и ретикулярный синцитий, сосредоточенный у основания жаберных лепестков), кишечник (слизистая), сердце (эпителиальный слой и эндотелий сосудов), почки (ретикулярный синцитий между канальцами), селезёнка, сосудистая кровь, лимфоидный орган (скопления кроветворной ткани – ретикулярного синцития – под крышей черепа). На отпечатках этих органов видны кровяные клетки разных стадий развития.
2. У костистых рыб наиболее активно гемопоэз происходит в лимфоидных органах, почке и селезенке, причем главным органом кроветворения являются почки (передняя часть). В почках и селезенке происходит как образование эритроцитов, лейкоцитов, тромбоцитов, так и распад эритроцитов.
3. Наличие в периферической крови рыб и зрелых и молодых эритроцитов является нормальным и не служит патологическим показателем в отличие от крови взрослых млекопитающих.
4. В эритроцитах, как и у других водных животных, в отличие от млекопитающих имеется ядро.
Селезёнка рыб располагается в передней части полости тела, между петлями кишечника, но независимо от него. Это плотное компактное тёмно-красное образование различной формы (шарообразной, лентовидной), но чаще вытянутой. Селезёнка быстро меняет объём под влиянием внешних условий и состояния рыбы. У карпа она увеличивается зимой, когда в связи с пониженным обменом веществ ток крови замедляется и она скапливается в селезенке, печени и почках, которые служат депо крови, тоже наблюдается при острых заболеваниях. При недостатке кислорода, перевозке и сортировке рыбы, облове прудов запасы крови из селезенки поступают в кровяное русло. Изменение размеров селезенки в связи с периодами усиленной активности установлено на ручьевой и радужной форелях и других рыбах.
Одним из важнейших факторов внутренней среды является осмотическое давление крови, так как от него зависит в значительной степени взаимодействие крови и клеток тела, водный обмен в организме и т. д.
Лимфатическая система рыб не имеет желез. Она представлена рядом парных и непарных лимфатических стволов, в которые лимфа собирается из органов и по ним же выводится в конечные участки вен, в частности в Кювьеровы протоки.
Дата добавления: 2015-06-22; просмотров: 860;