ЦЕСТОДОЗЫ
Гельминтозы, возбудителями которых являются представители класса ленточных червей Cestoidea, называются цестодозами. Класс ленточных червей включает в себя 9 отрядов, из которых в костистых пресноводных рыбах на взрослой фазе встречаются представители четырех отрядов: Caryophyllidea, Pseudophyllidea, Proteocephalidea и Nippotaeniidea. Представители четырех других отрядов (Trypanorhyncha, Diphyllidea, Tetraphyllidea и Lecanicepha-lidea) во взрослом состоянии паразитируют в акулах и скатах, а в костистых морских и проходных рыбах встречаются только их личинки — плероцеркоиды. Девятый отряд — Cyclophyllidea — объединяет высших цестод—паразитов теплокровных животных, у которых личиночные формы отдельных видов могут развиваться в различных внутренних органах пресноводных рыб. Среди ленточных червей далеко не все виды вызывают заболевания рыб. Однако есть виды весьма патогенные, вызывающие значительные эпизоотии, сопровождающиеся гибелью рыбы.
Тело ленточных червей молочно-белого цвета, плоское, лентовидное, состоит из головки (сколекса) и множества члеников (проглоттид), составляющих стробилу. Число члеников может достигать сотен и даже тысяч. У некоторых представителей этого класса, в частности отряда Caryophyllidea, тело не имеет члеников и выглядит как сплошная лента. Длина тела некоторых цестод может достигать 10—15 м. Половозрелые ленточные черви чаще всего паразитируют в кишечнике, личиночные стадии (плероцеркоиды) — в полости тела и других органах и тканях рыб.
Сколекс цестод служит для прикрепления и потому снабжен различными органами, приспособленными для этого: ботриями, ботридиями, присосками, хоботками. Прикрепительные органы надежно удерживают гельминта в кишечнике (рис. 77).
Наиболее примитивными органами прикрепления являются ботридии, которые представляют собой присасывательные ямки и бороздки на головке (1—4 у гвоздичников). Ботридии более разнообразны. Это — хорошо обособленные прикрепительные органы с развитой мускулатурой, располагающиеся на брюшной или спинной стороне сколекса — округлые, полые образования, снабженные собственной сильной мускулатурой, например у протеоцефалюсов. Хоботки — мышечные выросты, расположенные в верхней части головки; они могут втягиваться в специальное хоботное влагалище (четыреххоботники); хоботки обычно вооружены крючьями. Помимо хоботка хитиноидные крючья разнообразной формы и величины могут быть расположены на ботридиях (триенофорусы). За головкой следует шейка — зона роста, где формируются членики. Размер шейки различен. Молодые членики находятся сразу же за шейкой, старые — по мере роста червя отодвигаются все дальше от нее. В каждом членике имеется набор половых органов, которые развиваются в определенной последовательности: сначала закладываются элементы мужской половой системы, позднее женской, и членики становятся гермафродитнами. Последние членики стробилы почти целиком заполнены маткой, набитой яйцами.
Тело ленточных червей покрыто кутикулой, под которой находится кожно-мускульный слой. Полости тела у них нет и пространство между внутренними органами заполнено губчатой паренхиматозной тканью. Под кожно-мускульным слоем расположены продольные и поперечные мышцы.
Нервная система цестод представляет собой центральное нервное кольцо, располагающееся в головке. От него вперед и назад отходят продольные, нервные стволы, которые в каждом членике соединяются между собой поперечными кольцевыми комиссурами.
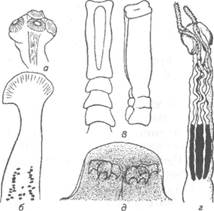
Рис. 77. Органы прикрепления цестод: а —присоски Silurotaenia siluri; б —ботрии
Caryophyllaeus laticeps; в — ботридии Bothriocephalus scorpii; г — хоботок Tetrarhynchus;
д— крючья Triaenophorus meridionalis червя (ботриоцефалюс). Присоски
Выделительная система состоит из мерцательных клеток, располагающихся в паренхиме. От них отходят мелкие канальцы, которые соединяются с выводными протоками (их 2 или 4). Они проходят в боковых частях члеников (соединяясь между собой комиссурами), в конце стробилы объединяются в экскреторный пузырек, открывающийся наружу. У нечленистых червей (гвоздичников) и ремнецов поперечные комиссуры отсутствуют и продольные выделительные протоки соединяются между собой беспорядочной сосудистой сетью.
Питание червей происходит путем всасывания всей поверхностью тела через кутикулу, на которой имеются микроскопические микротрихии, имеющие сложное строение, отличное от микро-ворсинок других плоских червей.
Половая система у расчлененных цестод самостоятельная в каждом членике, у нерасчлененных цестод имеется всего один половой комплекс. Все цестоды рыб — гермафродиты. Мужской половой аппарат состоит из семенников и выводных путей. Количество семенников различно: чаще их десятки, даже сотни, реже единицы. Они расположены в паренхиме, ближе к спинной стороне тела. От семенников отходят тонкие семявыносящие канальцы, которые сливаются в семяпровод, причем последний на конце может преобразовываться в семенной пузырек. Семяпровод идет к копулятивному органу — циррусу, способному выпячиваться; циррус расположен в мышечном мешке, называемом сумкой цир-руса. Входящая в сумку цирруса часть семяпровода является семя-извергательным каналом. Циррус и влагалище открываются в половую клоаку.
Женская половая система состоит из яичников, желточников, дополнительных желез, протоков и резервуаров, служащих для хранения и выведения половых продуктов.
В каждом половом комплексе имеется один, чаще двухлопастной яичник, от которого отходит яйцевод. В него открывается семяприемник и далее общий проток ведет к оотипу, в который поступают протоки желточников и железы Мелиса, способствующей образованию скорлуповой оболочки. В оотипе формируются яйца. Желточники чаще состоят из многочисленных фолликулов, которые располагаются в паренхиме, либо вокруг внутренних органов, либо в боковых частях члеников. От желточников отходят мелкие и тонкие желточные протоки, сливающиеся в два крупных протока, которые далее объединяются в один и открываются в оотип.
Вагина представляет собой трубку, связывающую женское половое отверстие с семяприемником, находящимся около оотипа. От оотипа берет начало матка, которая у многих цестод рыб имеет вид сильно извитой трубки, иногда мешковидной. Зрелые яйца выходят из матки в воду через ее отверстие, открывающееся самостоятельно на спинной или брюшной стороне членика или нерасчлененного червя. Помимо этого, зрелые яйца могут попадать в воду вместе с отторгнутыми члениками стробилы.
При совокуплении цестод циррус выпячивается и проникает в вагину этого же или соседнего членика или членика другой стробилы, лежащей рядом. Соответственно происходит самооплодотворение или перекрестное оплодотворение.
Эмбриональное развитие цестод рыб происходит в яйце, находящемся в большинстве случаев в матке, и попадающие в воду зрелые яйца содержат сформированных эмбрионов, имеющих 6 крючьев (онкосфера) или 10 крючьев (ликофора). В воде из яйца выходит личинка, называемая корацидием, которая некоторое время ведет свободный образ жизни. Тело корацидия покрыто ресничками, имеет 3 пары подвижных эмбриональных крючьев. У некоторых цестод личинки не выходят из яйца, а попадают в промежуточного хозяина при поедании им зрелых яиц паразита.
Дальнейшее развитие цестод происходит с одной или двумя сменами промежуточных хозяев. Первым промежуточным хозяином цестод рыб обычно являются низшие ракообразные (чаще всего веслоногие), бокоплавы, мизиды, реже малощетинковые черви (у гвоздичников) и другие беспозвоночные.
Яйца с онкосферой (у гвоздичников) или свободноплавающие корацидии (у ремнецов, ботриоцефалюса, триенофоруса и др.) заглатываются беспозвоночными животными, сквозь стенки их кишечника проникают в полость тела, превращаясь в процеркои-да — первую паразитическую стадию цестод.
Процеркоид в полости тела промежуточного хозяина увеличивается в размерах; развиваются его внутренние системы (мышечная ткань, выделительная система). На заднем конце его образуется особый придаток — церкомер, в котором находятся зародышевые крючья.
Дальнейшее развитие цестод различается в зависимости от того, с одним или с двумя промежуточными хозяевами протекает жизненный цикл гельминта. Если цикл развития происходит при участии одного промежуточного хозяина (гвоздичники, ботриоце-фалюс, протеоцефалюс), то уже у процеркоида формируются прикрепительные органы, характерные для взрослых червей, закладывается и развивается половая система. Попадая вместе с пищей в кишечник окончательного хозяина, процеркоид прикрепляется к стенкам кишечника и развивается во взрослого червя.
Если цикл развития происходит при участии двух и более промежуточных хозяев, то процеркоид вместе с первым промежуточным хозяином (ракообразным) попадает в кишечник второго промежуточного хозяина, проникает в полость его тела (ремнецы), печень (триенофорус), мускулатуру (лентец широкий), где теряет церкомер и превращается в следующую личиночную стадию — плероцеркоид.
Плероцеркоид растет. У ремнецов развиваются внутренние органы, заканчивается органогенез половой системы, у лентецов плероцеркоид слабо дифференцирован и внутренние органы его еще не заложены. Инвазионного состояния плероцеркоиды ремнецов достигают через 6—14 мес. Далее плероцеркоид вместе со вторым промежуточным хозяином попадает в кишечник окончательного хозяина, где достигает половой зрелости и продуцирует яйца.
Кавиоз
Возбудитель заболевания — цестода Khawia sinensis — относится к отряду гвоздичников Caryophyllidea, сем. Lytocestidae. Паразитирует в кишечнике карпа, сазана и их гибридов. Цестоды завезены в европейскую часть России из р. Амур вместе с амурским сазаном.
Возбудитель. Тело гельминта нечленистое, белое, длиной 80—170 мм и шириной 3,5—5,0 мм. Головной конец паразита веерообразно расширен, с фестончатым передним краем. Шейки нет. Семенники и желточники расположены, несколько отступя от головы. Имеется один половой комплекс, в котором находятся единичные желточные фолликулы; в заднем конце тела имеется большая постовариальная группа желточников. Яичник Н-образный, передние лопасти его значительно шире задних. Петли матки не заходят вперед сумки цирруса (рис. 78, а).
Развитие К. sinensis происходит при участии одного промежуточного хозяина — мало-щетинковых червей (олигохет) (рис. 79). С экскрементами рыбы яйца гвоздичников (чаще в апреле—мае) попадают на дно водоема, где в течение 30—40 дней происходит развитие зародыша. Сформировавшийся в яйце корацидий не выходит из яйца, а находится в нем до тех пор, пока яйцо проглотит промежуточный хозяин — олигохета (Tubifex tubifex, Limnodrilus udeke-mianus и др.). В нем онкосфера освобождается от оболочки яйца и проникает в полость тела червя, где в передней его части происходят формирование церкомера и рост паразита. Эта личиночная стадия носит название процеркоида. Длина его достигает 1,5—2,0 мм. Процеркоиды могут вызывать кастрацию и даже гибель трубочников.
Развитие процеркоида в трубочнике продолжается 2—3 мес (весна—лето). Когда рыба съедает трубочника, вместе с ним про-церкоид попадает в ее кишечник, где становится половозрелым. К. sinensis могут перезимовывать в рыбе и олигохетах.

Рис. 78. Возбудители кавиоза и кариофилеза: a —Khawia sinensis; б — Caryophyllaeus fimbriceps; I vi II — передний и задний концы цестод; 1 — сколекс; 2 —желточники; 3 —семенники; 4 — сумка Цирруса; 5—матка; 6 —яичник; 7—половая бурса

Рис. 79. Цикл развития гвоздичника
Эпизоотология. Заболеванию подвержены двухлетки и сеголетки карпа, сазана и сазано-карповых гибридов, которых сейчас разводят в прудовых хозяйствах страны. В последние годы К. sinensis стали иногда обнаруживать у сеголетков и годовиков черного и белого амуров. Старшие возрастные группы рыб болеют редко, но являются паразитоносителями.
Гвоздичник К. sinensis распространен довольно широко в рыбоводных хозяйствах России, стран Балтии, Беларуси, Украины, государств Средней Азии.
Вспышки заболевания в прудовых хозяйствах отмечаются чаще всего в мае—июне, когда зараженность двухлетков карпа достигает 40 % ( к середине мая), а в июне — 80 %. К концу августа зараженность обычно снижается до 20—30 %, уменьшается и интенсивность поражения. Зимой у карпов встречается небольшое количество молодых гвоздичников. Наиболее опасно заболевание для сеголетков карпа, у которых оно вызывает снижение темпа роста.
Гибель рыбы может происходить только при поражении несколькими десятками паразитов. Было отмечено заболевание среди трехлетков карпа, в кишечнике которых паразитировало до 45 крупных гвоздичников.
Источником инвазии служат зараженные рыбы и инвазирован-ные трубочники. Последние весьма стойки в отношении различных дезинфектантов, а при недостаточном осушении прудов могут выживать благодаря способности мигрировать глубоко в грунт.
Клинические признаки и патогенез. Больные рыбы плавают у поверхности, отказываются от корма. При высокой интенсивности инвазии черви закупоривают кишечник, нарушая процесс пищеварения.
Диагноз. Его ставят на основании эпизоотологических данных, клинического и паразитологического анализа, при котором определяются видовая принадлежность и численность гельминта.
Меры борьбы. На неблагополучное хозяйство накладывают ограничения и проводят комплекс ветеринарно-санитарных мероприятий. Пруды, в которых содержалась инвазированная рыба, тщательно просушивают весной перед зарыблением и осенью после отлова рыбы. Зимой пруды промораживают с дезинфицирующей обработкой ложа негашеной известью из расчета 2500— 3000 кг/га, или хлорной известью — 500 кг/га. Проводят поочередное летование прудов с перепахиванием ложа и посевом трав.
В некоторых случаях (при высокой степени зараженности, затруднениях при проведении обычных мероприятий по снижению численности олигохет) рекомендуется разводить рыб других видов, питающихся олигохетами, но не заражающихся кавиями (линь, карась).
Для дегельминтизации рыб используют гранулированные лечебные корма с фенасалом или с микросалом (циприноцестин-2). Лечебное кормление проводят в течение одного дня без предварительной голодной диеты при суточной дозе, соответствующей обычному кормлению рыбы комбикормом.
При отсутствии готовых лечебных кормов проводят одноразовую дегельминтизацию фенасалом или микросалом, вводя их 1 % или 2 % соответственно к задаваемой суточной норме корма. Производителей и ремонтную группу рыб можно дегельминтизиро-вать индивидуально. Для этого водную суспензию фенасала вводят при помощи шприца и мягкого резинового катетера из расчета 0,5 г препарата на 1 рыбу массой 0,5—1,5 кг, но не более 1 г на одного производителя.
В качестве других лечебных препаратов можно использовать камалу, горчицу, табачную пыль, порошок из высушенных корневищ папоротника (щитовника шиповатого).
Дата добавления: 2015-06-22; просмотров: 1717;