СПОСОБНОСТЬ К ОБУЧЕНИЮ
Способность к обучению – это наиболее трудная, но и наиболее увлекательная область генетики поведения. Первая последовательная попытка селекции крыс на способность к обучению (опыт Трайона) показала практически все трудности, подстерегающие исследователя на этом пути.
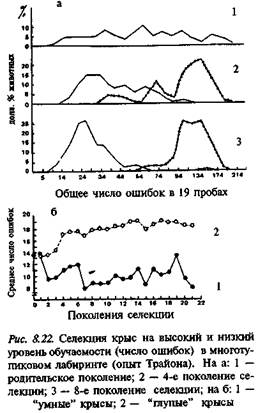
8.6.3.1. линии ТРАЙОНА. Селекция крыс на успешность (количество ошибочных заходов в тупики) обучения поиску пищи в лабиринте (рис. 8.22) была одной из первых работ по генетике поведения (см.: 8.6.2). Было показано, что крысы "умной" линии ТМВ обучались значительно лучше в исходном 17-тупиковом лабиринте, а также в более простом, 14-тупиковом. В то же время при обучении в 16- и б-тупиковых лабиринтах межлинейных различий обнаружено не было.
Обучение реакции избегания воды было более успешным у "глупой" линии TMD. В дальнейшем было показано, что ТМВ лучше обучались реакции активного избегания в челночной камере, но в тесте на обучение избеганию тока, когда надо было выпрыгивать из ры (а не переходить из одной ее половины в другую) показатели TMD были выше. Это может означать, что в эксперименте Трайона отбор животных производился (неосознанно, разумеется) не только на способность к обучению, но и на какие-то особенности поведения, связанные с лабиринтом данной конфигурации. У крыс линии ТМВ выше пищевая мотивация, но слабее оборонительная, они меньше отвлекаются при выполнении навыка, тогда как крысы линии TMD пугаются даже при манипуляциях дверцами лабиринта. Исследователи приходят к выводу, что ТМВ лучше решают тесты, связанные с пространственными стимулами, тогда как TMD успешнее обучаются при зрительных раздражителях.
|
Другие формы поведения трайоновских линий в значительной степени различаются между собой, причем физиологическая интерпретация различий иногда достаточно проста, иногда затруднительна или просто невозможна. У линии ТМВ был ниже порог провокации ЭЭГ-знаков судорожной активности при электростимуляции миндалины, слабее реакция на новые предметы, ниже уровень локомоторной активности в колесах. Можно полагать, что отбор на разную степень успеха обучения в лабиринте способствовал формированию генотипов, при которых особенности процесса восприятия, параметры пространственной памяти, мотивация и другие феноти-пические признаки оказались в одном случай оптимальны, а в другом – субоптимальны для выполнения данной реакции.
Результаты сравнения поведения трайоновских линий достаточно поучительны в том отношении, что иллюстрируют сложную картину, которая может получиться в результате отбора на способность к обучению, если реально отбору подвергается признак, косвенно  связанный с изучаемым. Несомненно, что при создании путем отбора трайоновских линий произошло видоизменение их поведенческих фенотипов. В то же время достоверного прямого влияния отбора на собственно способность к обучению не обнаруживается. Примером сложности полученной картины является изменение способности к обучению крыс этих линий при выращивании их в условиях с разной степенью "обогащенности" среды (рис. 8.23; см. также: 7.3).
связанный с изучаемым. Несомненно, что при создании путем отбора трайоновских линий произошло видоизменение их поведенческих фенотипов. В то же время достоверного прямого влияния отбора на собственно способность к обучению не обнаруживается. Примером сложности полученной картины является изменение способности к обучению крыс этих линий при выращивании их в условиях с разной степенью "обогащенности" среды (рис. 8.23; см. также: 7.3).
8.6.3.2. ГЕНЕТИЧЕСКИЕ ИССЛЕДОВАНИЯ УСЛОВНОЙ РЕАКЦИИ АКТИВНОГО ИЗБЕГАНИЯ. Реакция активного избегания удара электрического тока в челночной камере – это четкий, легко поддающийся количественному учету тест на обучаемость у лабораторных грызунов. Поскольку в качестве условного раздражителя можно выбрать звук, то при использовании этого теста можно включать в сравнение и животных-альбиносов, не опасаясь, что низкая острота их зрения повлияет на успешность выполнения навыка. Челночные камеры, в которых животные обучаются попеременно переходить из одного отсека в другой при предъявлении условного сигнала, легко автоматизировать, что способствует получению данных на большом числе животных в унифицированных условиях. Все это сделало данную методику очень популярной среди исследователей, а число работ, выполненных с ее помощью, составляет многие сотни. Особенно велико число фармакологических и фармакогенетических исследований.
В начале 60-х годов на основе популяции крыс Вистар итальянский исследователь Дж. Биньями начал селекцию на высокую и низкую способность крыс к обучению реакции активного избегания в челночной камере. Уже через несколько лет эти линии были сформированы. С начала 70-х годов селекция и разведение этих крыс проводились независимо в Швейцарии, Великобритании и Канаде.
Межлинейные различия по селектируемому признаку высокодостоверны. Эти линии получили название римских (Roman High Avoidance, RHA, Roman Low Avoidance, RLA) и интенсивно исследуются во многих лабораториях мира.
Сравнение результатов огромного числа работ, полученных на этих линиях с использованием практически всех существующих методик оценки поведения, привело исследователей к заключению, что межлинейные различия у RHA и RLA связаны с эмоциональностью и разным типом реакции на стрессоры (Жуков, 1997; см. также: 8.6.5.3). В то же время существуют достаточно убедительные данные, что эти линии показывают различия и в ассоциативных способностях, т.е. в способности к обучению как таковой. Большое внимание нейроэндокринологическим механизмам различных стратегий поведения крыс в условиях избегаемого и неизбегаемого удара электрического тока было уделено в работах на крысах двух селектированных линий – КНА и KLA (Koltoushi High Avoidance, Koltoushi Low Avoidance), иначе называемых линиями BJI и НЛ. Выведение этих линий было начато в конце 50-х годов в Институте физиологии им. Павлова АН СССР В.К. Федоровым, а после его смерти продолжено Н.Г. Лопатиной с соавт. (см.: Лопатина, По-номаренко, 1987). Как и другие линии крыс, селектированные на высокую и низкую способность к избеганию в челночной камере, данные линии отличаются друг от друга и по иным признакам, связанным как с поведением, так и с более "простыми" физиологическими реакциями. Подробный анализ экспериментальных данных, полученных на крысах этих линий, проведен в монографии ДА. Жукова (1997).
Способность крыс селектированных линий к обучению реакции избегания была сопоставлена с таким морфологическим показателем, как размер интра- и инфрапирамидных проекций мшистых волокон – аксонных окончаний нейронов зубчатой фасции в поле САЗ гиппокампа. Эта проблема рассмотрена более подробно в разделе 8.6.7.2.
Позднее, уже в 70-е годы были созданы путем селекции еще две линии (Syracuse High Avoidance, SHA, Syracuse Low Avoidance, SLA), различающиеся по обучению реакции активного избегания, но не обнаруживающие, в отличие от римских линий, различий в уровне двигательной активности. Исходной популяцией для этой селекционной работы служили крысы Лонг-Иване. В настоящее время крысы хорошо обучающейся линии SHA дают примерно 40 реакций избегания в 60 предъявлениях теста, тогда как SLA – ни одного. У SLA сильно подавлены межсигнальные реакции, однако скорость выполнения реакций избавления (т.е. реагирования на включение электрического тока-наказания) у них не отличалась от линии SHA. В тесте "открытого поля" SLA, как и RLA, более эмоциональны, однако различий в уровнях локомоции у них, как сказано выше, не выявлено.
Генетические модели способности к обучению, какими являются линии крыс RHA и RLA, а также SHA и SLA, используются также в исследованиях роли наиболее изученных медиаторных систем в формировании межлинейных различий реакции на стресс, в обучении, в организации болевой чувствительности. Эти исследования представляют большой интерес, но в настоящем пособии не могут быть затронуты сколько-нибудь подробно (см.: Gray, 1987).
Другим аспектом проблемы, который делается очевидным при анализе межлинейных различий в обучении реакции избегания в челночной камере, является биологическая адекватность самого теста. В самом деле, анализ "логики" этого теста показывает, что данная реакция далека от того, чтобы имитировать какую-либо из естественных поведенческих реакций крысы или мыши. Животное, находящееся в камере, получает болевое раздражение (удар тока), которому предшествует некий сигнал (свет и/или звук). Бурная двигательная реакция в ответ на боль "переносит" крысу или мышь в другую половину камеры, где ток (и наказание) отсутствует и где она получает отдых. Однако через короткое время в этом бывшем безопасном отсеке животное получает новый удар тока, а метания по клетке переносят его в отсек, где ранее также предъявлялось наказание. В этих условиях у животных (тем не менее) формируется условная реакция выполнения перехода из одного отсека в другой в ответ на предъявление условного сигнала (света или звука).
Реакция перехода выполняется в двух половинах камеры (в двух разных контекстах), причем пребывание в каждом из них регулярно наказывается (если реакция не выполняется). Это означает, что в данном тесте животное должно искать убежища там, где оно только что получило наказание.
Возможно, эта логическая противоречивость теста и есть причина того, что множество генетических линий и групп лабораторных грызунов демонстрирует весьма невысокие показатели обучаемости, хотя у некоторых животных они, напротив, высокие. В чем же заключаются различия между такими животными? Ответ на этот вопрос исследователи нашли с помощью генетико-этологического анализа проблемы (Вилкок, 1975). Генетический эксперимент, который позволил найти путь его разрешения, изложен в разделе, посвященном описанию методов диаллельного скрещивания (см.: 8.6.6.2).
Этологический аспект проблемы заключается в следующем. Боль как фактор, провоцирующий у животного реакцию страха, вызывает у млекопитающих, а точнее у грызунов, одну из двух ответных реакций: либо замирание (затаивание), либо бегство.
Существует так называемая двухкомпонентная теория физиологических механизмов реакции избегания, в соответствии с которой в начальный период обучения выявляется тенденция либо "затаиваться" от опасности, которую представляет собой боль, предваряемая условным сигналом, либо убегать, но обе реакции сначала осуществляются вне связи с условным сигналом. Преобладание у животных реакции "затаивания" ухудшает их шанс усвоить данный навык. В целом, по мере увеличения числа сочетаний условного и безусловного сигналов в поведении начинает превалировать тенденция к бегству. Это не объясняет, почему смена реакций обнаруживает подобную динамику, однако для анализа поведения в челночной камере с позиций генетической изменчивости объяснение представляется продуктивным.
При отборе животных в процессе выведения линий, быстро и медленно обучающихся условнорефлекторной реакции избегания, исследователь руководствуется условными критериями обученное™, точнее, тем, как быстро животные, которые будут использованы в качестве производителей, их достигают. Можно предполагать, что крысы, быстрее достигшие данного критерия, действительно обладают более высокими ассоциативными способностями. Однако естественно предположить и другое: исследователи отбирают не только "умных" и "глупых" животных, но и таких, у кого оказываются более низкими пороги реакции бегства в ответ на боль (более высокий уровень реакций избегания), а также тех, у кого ниже пороги реакции замирания как ответа на боль (уровень реакций избегания ниже). Данные опыта по диаллельному скрещиванию (см.: 8.6.6.2) в целом подтверждают правомочность такого предположения.
Дата добавления: 2015-01-10; просмотров: 1759;