ПЕНТОЗОФОСФАТНЫЙ ПУТЬ
Пентозофосфатный путь (ПФП) обмена углеводов нередко называют апотомическим путём, так как обмен глюкозы идёт по первому (С1) атому углерода.
Доля ПФП в количественном превращении глюкозы в клетках обычно невелика (в большинстве клеток не более 10 %) и варьирует в зависимости от типа ткани и её функ-ционального состояния. Так, в клетках печени по этому пути превращается до 20 % глюкозы, в эритроцитах — 7 %, в клетках мозга — около 2 %. Этот процесс идет в клетках многих органов и тканей.
Ферменты ПФП локализованы в цитоплазме клеток.
Превращение глюкозы по ПФП не требует присутствия кислорода. Если по ПФП превращается шесть молекул Гл-6-Ф, то за один цикл молекула Гл-6-Ф катаболизирует до 6 СО2.
Суммарное уравнение:
6 Гл-6-Ф + 7H2O + 12 НАДФ+ ® 5 Гл-6-Ф + 6СО2 + 12 НАДФН.Н+ + ФН
Последовательность реакций ПФП разделяют на два этапа:
I. Окислительный этап. На этом этапе осуществляются две дегидрогеназные реакции и одна реакция декарбоксилирования с образованием рибозо-5-фосфата и восстановлением двух молекул НАДФ+ (2 НАДФ+ → 2 НАДФН.Н+)
Реакции:
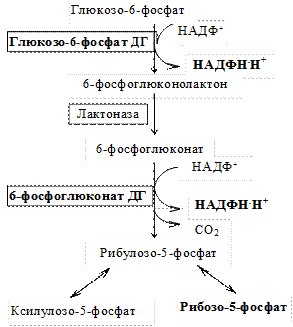
Таким образом, при окислении молекулы глюкозы образуется 2 НАДФН.Н+ и рибозо-5-фосфат. В некоторых клетках катаболизм глюкозы на этом и заканчивается.
Ключевые ферменты:
1)глюкозо-6-фосфатдегидрогеназа — главный ключевой фермент;
2) 6-фосфоглюконатдегидрогеназа.
Значение окислительного этапа:
1. Главный поставщик рибозо-5-фосфата для биосинтетических процессов:
- биосинтез мононуклеотидов (АМФ, ГМФ, УМФ, ЦМФ, ТМФ и др.);
- синтез нуклеиновых кислот (ДНК, РНК);
- синтез коферментов (НАД+, НАДФ+, ФАД, КоА-SН).
2. Основной источник НАДФН.Н+ в клетках. ПФП на 50 % обеспечивает потребности клетки в НАДФН.Н+.
НАДФН.Н+ в клетках используется:
1) в реакциях биосинтеза веществ как восстановитель:
- синтез жирных кислот;
- биосинтез холестерола, стероидных гормонов, желчных кислот;
- синтез заменимых аминокислот (НАДФН·Н+ как кофермент глутаматдегидрогеназы в реакциях восстановительного аминирования α-кетоглутаровой кислоты);
- в глюкуроновом пути и др.
2) в обезвреживании веществ: в реакциях гидроксилирования различных ксенобиотиков, лекарственных веществ, этанола и других веществ, которые осуществляются с участием микросомной цитР450-зависимой системы окисления;
3) как антиоксидант: используется на восстановление окисленного глутатиона. Глутатион — важного антиоксиданта клеток;
4) в фагоцитозе: генерирование активных форм кислорода. Фагоциты с использованием НАДФН.Н+ генерируют супероксидные анион-радикалы, выполняющие основную роль в разрушении поглощённых бактериальных клеток. При недостаточной продукции НАДФН.Н+ при нарушении ПФП отмечается хроническое течение инфекционных заболеваний.
Интенсивность протекания реакций ПФП зависит от потребности клеток в продуктах реакций и различается в разных тканях. Реакции окислительного этапа активно протекают в клетках печени, жировой ткани, эмбриональной ткани, в коре надпочечников, щитовидной железе, половых железах, лактирующей молочной железе, костном мозге, эритроцитах.
II. Неокислительный этап (этап межмолекулярных перегруппировок). На этом этапе происходят взаимопревращения сахаров (фосфотриоз, фосфотетроз, фосфопентоз, фосфогексоз, фосфогептулоз, фосфооктулоз), в результате которых регенерирует глюкозо-6-фосфат.
Два основных фермента катализируют превращения на неокислительном этапе:
1)транскетолазакатализирует перенос двухуглеродных фрагментов. В качестве кофермента использует тиаминпирофосфат;
2) трансальдолазакатализирует перенос трёхуглеродных фрагментов.
Варианты неокислительных превращений:
· классический или F-вариант (от англ. fat — жир) — осуществляется в клетках жировой ткани;
· октулозный или L-вариант (от англ. liver — печень) — осуществляется в клетках печени и других тканей.
Реакции (L-вариант):
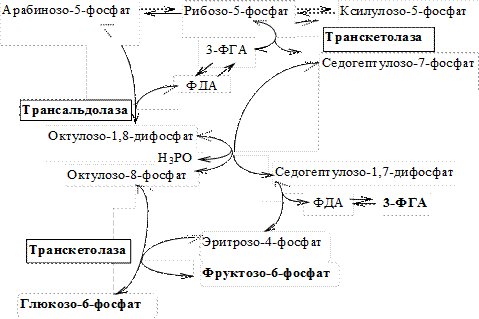
Итак, на неокислительном этапе невостребованные в клетках пентозофосфаты в результате межмолекулярных перегруппировок превращаются в Гл-6-Ф, а также образуются Фр-6-Ф и 3-ФГА.
Все реакции неокислительного этапа обратимы.
На неокислительном этапе ПФП связан с гликолизом (посредством Гл-6-Ф, Фр-6-Ф и 3-ФГА), то есть возможно переключение этих процессов.
Значение неокислительного этапа:
1. Стабилизирует концентрацию фосфопентоз в клетке, то есть утилизирует лишние фосфопентозы. Благодаря связи с гликолизом лишние пентозы катаболизируют по гликолитическому пути, давая клеткам энергию.
2. Синтез фосфопентоз в клетке при торможении окислительного этапа благодаря обратимости реакций неокислительного превращения.
Регуляция пентозофосфатного пути, в основном, осуществляется на уровне дегидрогеназ. Инсулин индуцирует синтез глюкозо-6-фосфатдегидрогеназы, 6-фосфоглюконат-дегидрогеназы. Жирные кислоты — аллостерические ингибиторы глюкозо-6-фосфат-дегидрогеназы. Увеличение уровня НАДФН.Н+ в клетке тормозит окисление глюкозы по ПФП.
Дата добавления: 2015-03-11; просмотров: 1121;