Глава 11. ВОСПРОИЗВОДСТВО РЫБ
По биологическому значению функция размножения у животных уступает только добыванию корма. Однако в период высокой половой активности половая доминанта подавляет и чувство голода. В водной среде процесс размножения отличается относительной простотой по технике исполнения. Чаще всего оплодотворение в воде носит вероятностный характер, а потому количество вариантов должно быть достаточно большим, чтобы сделать процесс оплодотворения результативным. Поэтому в целом для класса рыб характерна высокая плодовитость. Количество женских гамет за один период икрометания у представителей пелагических видов рыб достигает десятков и даже сотен миллионов, а количество сперматозоидов и того больше.
Среди животных с таким внутриклассовым морфофункциональным разнообразием, как рыбы, всегда есть исключения из общего правила. Это справедливо и по отношению к процессу размножения.
Существует ряд общих требований, без которых размножение невозможно. Во-первых, рыбы должны достичь возраста полового созревания. Правда, у рыб понятие "возраст полового созревания" достаточно расплывчато и не так привязано к календарному возрасту, как у высших позвоночных.
Межвидовые различия сроков полового созревания у рыб огромны: от 1 мес у гамбузии до 30 лет у некоторых видов осетровых рыб. У рыб сроки полового созревания помимо календарного возраста определяет ряд факторов внешней среды (температура воды, химический состав воды, интенсивность питания, плотность посадки в искусственные водоемы). Время полового созревания коррелирует с живой массой рыб.
Сроки полового созревания различаются у рыб разных популяций одного вида. Например, у леща половая зрелость наступает в популяции Ладожского озера в 8-9 лет, в популяции леща Средней Волги -в 6-7 лет, в Северном Каспии - в 4-6 лет. Азово-черноморская популяция леща начинает нереститься уже в 3 года.
То же можно сказать и о других видах рыб. Например, у волжского и азовского осетров половая зрелость наступает в 9-10 лет. У куринского осетра половая зрелость самок наступает лишь в 30 летнем возрасте.
§51. ДИФФЕРЕНЦИАЦИЯ ПОЛОВ
Половая дифференциация у рыб - явление уникальное. Рыбы обладают очень пластичной системой репродукции, которая позволила им занять практически все существующие на Земле водоемы. В соответствии с биологической необходимостью рыбы могут изменять половую принадлежность отдельных особей. У рыб нет единообразной хромосомной системы полового детерминизма, как, например, у человека (XX-, XY-хромосомы. т. е. системы половых гетерохромосом). Точнее, как вариант она встречается в классе рыб, например у рыбца. Самки рыбца в диплоидном наборе имеют две одинаковые половые хромосомы (XX), самцы рыбца имеют две разные половые хромосомы (XY).
У других видов, например японского угря, картина прямо противоположная: у самца имеется пара одинаковых половых хромосом (ZZ), а у самки -две разные половые хромосомы (WZ). Кроме того, есть виды, у которых у одного из полов половая хромосома непарная. Так, например, самка фундулюса имеет парные половые хромосомы (XX), а самец - непарную половую хромосому (ХО).
У лабиринтовой рыбы коллизии только самец несет половую хромосому, да и ту непарную.
У основных объектов рыборазведения -- карповых, лососевых, представителей осетровых рыб -половых хромосом нет. У этих рыб половой детерминизм имеет полихромосомную основу, т. е. гены, кодирующие первичные и вторичные половые признаки, рассредоточены по другим соматическим хромосомам.
Такие резкие различия полового генно-хромосомного детерминизма у рыб едва ли связаны с филогенезом, так как они встречаются в пределах одного семейства и даже в пределах одного вида (последнее характерно для разных популяций угрей и пециллий).
Неопределенность с половой принадлежностью остается и в онтогенезе. Чаще всего у молоди нет половых различий до момента полового созревания. При этом даже гистологические исследования гонад не проясняют ситуации с полом.
У рыб первичные половые клетки способны развиваться как по Женскому, так и по мужскому пути. Многим видам рыб свойствен ювенальный гермафродитизм, т. е. параллельное развитие и мужских, и женских гонад и соответственно половых клеток, одни из которых впоследствии отмирают.
У рыб известно явление проандрии и прототинии, когда на раннем этапе онтогенеза рыбы развиваются мужские гонады, а на Последующем - женские. Как нормальное физиологическое явление у рыб встречается несколько типов функционального гермафродитизма. Особенно много примеров этого явления у окуневых рыб. Есть виды окуней, которые первую половину репродуктивного периода являются самками, а вторую- самцами. При этом рыбы имеют и первичные, и вторичные половые признаки с нормальным оно- или сперматогенезом и соответствующим полу нерестовым поведением.
И уж совсем необычно выглядит истинный суточный транссексуализм у морских окуней. В течение суток одна и та же особь многократно меняет свою половую принадлежность, выметывает или икру, или молоки. Самооплодотворение здесь маловероятно, но возможно.
Вторичные половые признаки рыб, которые особенно ярко проявляются в период нереста, имеют важное практическое значение в рыбоводстве при сортировке рыб по половому признаку.
Например, у лососей признаком самцов являются более яркая окраска и изменения опорно-двигательного аппарата - искривление челюстей, появление горба.
У самца колюшки перед нерестом брюшко окрашивается в алый цвет. Самцы многих морских бычков перед нерестом становятся абсолютно черными.
У карповых (вобла, язь) на голове и теле самцов в преднерестовый период появляется "жемчужная сыпь" - роговые образования белесого цвета.
Появление брачного наряда у рыб определяется гормональными перестройками в этот период жизни. Значение его специалисты трактуют по-разному. Брачная окраска в нерестовый период может выполнять защитную роль. У лососевых брачный наряд маскирует рыб, делает их менее заметными на галечном грунте в прозрачной воде. В других случаях нерестовая окраска имеет сигнальное значение (горбуша, кета).
Пол одних рыб можно определить по форме анального отверстия, других - по форме плавников (рис. 11.1).
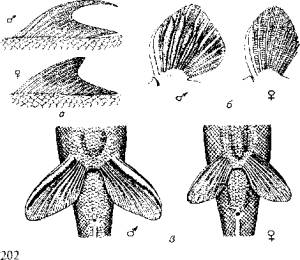
Рис. 11.1. Вторичные половые признаки рыб:
а-спинные плавники Lafaeo dero; б-грудные плавники тибетского гольца; в - брюшные плавники линя
Например, у живородящих пициллид половой диморфизм довольно рано проявляется в особом строении анального плавника самцов, выполняющего роль совокупительного органа. У акул такую же роль выполняет придаток брюшного плавника.
Важным половым признаком может служить половое поведение рыб перед нерестом. Это может быть преследование самцом самки (карп, щука и др.) или более сложное поведение - устройство гнезда (лабиринтовые), охрана территории. После нереста у многих видов рыб наблюдаются элементы родительского поведения: вентиляция икры, инкубация икры в ротовой полости, охрана молоди (тиляпия).
Конечно, какими бы сложными формами поведения рыбы не обладали в период размножения, все они являются результатом врожденных стереотипических поведенческих актов. Половое поведение проявляется при изменении гормонального статуса рыб в ответ на действие внешней среды. Половая доминанта формируется под влиянием усиления функции гипоталамо-гипофизарно-гонадо-адреналовой системы.
Управление половым детерминизмом при искусственном разведении рыб имеет большое практическое значение. Особенно полезным этот прием может быть при разведении ценных рыб - осетровых, лососевых. Здесь желательно иметь большое поголовье самок и ограниченное количество самцов, однако половые признаки у этих видов проявляются довольно поздно, когда уже затрачено много средств и времени.
Регулировать пол в искусственных условиях выращивания удается при помощи стероидных гормонов. Андрогены и эстрогены не разрушаются в желудочно-кишечном тракте. Поэтому они добавляются в корма. Включения метилтестостерона в рацион личинок тиляпии в количестве 30-50 мг/кг приводит к тому, что в стаде половозрелых рыб самцы составляют 95-100 %. При добавлении в рацион форели этого же гормона (3 мг/кг) все особи превращались в самцов.
При добавлении в рацион гормона эстрадиола (20 мг/кг корма) у лососей формировалось полностью (на 100 %) женское гомосексуальное стадо. Такой же "феминистический" эффект получали у тиляпии при помощи этинилэстрадиола в количестве 50 мг/кг корма.
Таким образом, гормональная регуляция пола у рыб при помощи синтетических или природных стероидов может претендовать на самостоятельный технологический прием в рыбоводстве.
§52. ОСОБЕННОСТИ ОВОГЕНЕЗА И СПЕРМАТОГЕНЕЗА
К моменту полового созревания рыб масса их гонад (семенников и ястыков) возрастает в результате деления половых клеток. До определенного момента гонады самцов и самок не различаются по массе. Однако в последний период перед нерестом происходит активное накопление питательных веществ в ястыках и поэтому разница в массе ястыков и семенников перед нерестом становится весьма существенной (рис. 11.2).

Рис. 11.2. Изменение массы гонад у форели в течение года (г/кг массы тела)
У некоторых рыб с внутренним оплодотворением бывает асинхронное созревание самок и самцов. В этом случае совокупление приводит к тому, что сперма хранится в половых путях самки до времени овуляции, когда и происходит оплодотворение икринок.
Чаще масса икры превосходит массу спермы при одинаковой живой массе самцов и самок (табл. 11,1).
В тропическом поясе у многократно нерестующих рыб масса выметанной икры может превысить массу тела самой самки.
Не все половые клетки достигают полного развития и резорбируются. Компоненты этих клеток служат питательным материалом для дальнейшего развития других зигот.
Половые клетки богаты белками, жирами и биологически активными веществами, такими, как витамины, гормоны, простагландины.
Морфологически половая система рыб очень разнообразна и тем не менее достаточно проста (рис. 11.3). У рыб семенник соединен с мочеточником. Сперма извергается по мочевому каналу в клоаку.
У самок после овуляции икринки попадают в брюшную полость, затем захватываются воронкой яйцевода и выводятся в клоаку.
У круглоротых процесс упрощен. Сперма и икра при созревании гонад поступают в брюшную полость. Затем половые клетки через поры проникают в просвет мочеточника.
У осетровых и акул мочевые протоки головной почки формируют мюллеровы протоки, по которым половые продукты и самца, и самки после созревания покидают половые железы.
Овуляция и эякуляция находятся под гормональным контролем. Овуляция протекает сразу во всем ястыке и захватывает все зрелые овоциты, созревание которых происходит циклично. Для самок характерен порциальный нерест.
11.1. Масса зрелых гонад, %
| Вид рыб | Самки | Самцы |
| Белый толстолобик | 10-15 | 0,4-0,5 |
| Карп | 10-25 | 2-9 |
| Радужная форель | 10-15 | 5-8 |
| Треска | До 30 | До 30 |

Рис. 11.3. Строение половой системы рыб:
А-минога; б- акула; в- осетр; г - лосось; д - карп; 1- яичник; 2 - яйцевод: 3 - вторичный яйцевод; 4 - семенник; 1 - семяпровод; 6- почка; 7- мочеточник; 8 - клоака; 9 - мочеполовой синус
Сперматогенез - процесс более равномерный, чем овогенез. Поэтому самцы участвуют Б нересте более продолжительное время.
Плодовитость рыб связана с величиной икринок. Так, у акулы диаметр яйца достигает нескольких сантиметров, причем самки могут вынашивать всего одно яйцо. У костистых рыб при величине икринки менее 1 мм их общее количество достигает нескольких миллионов (табл. 11.2).
Дата добавления: 2014-12-22; просмотров: 2742;