Предмет ГП как отрасли права.
В хлоропласте осуществляется первичное взаимодействие света с пигментом, происходит синтез углеводов клетки из СО2 и Н2О и синтезируется АТФ в процессе фотофосфорилирования.
Взаимодействие света с пигментами, синтез АТФ и восстановление НАДФ+ происходят в тилакоидах хлоропластов. В строме органоида осуществляются реакции цикла Кальвина и синтез крахмала из триозофосфатов через превращение части их в гексозофосфаты. Не использованные на синтез крахмала триозофосфаты потребляются на общие нужды клетки. Синтезированные в тилакоидах АТФ и НАДФ, а также поступающий извне СО2 участвуют в строме в цикле Кальвина.
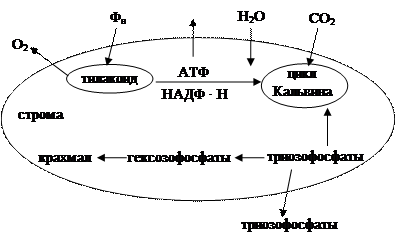 |
Таким образом, хлоропласт, получая извне СО2, Н2О и неорганический фосфат, поставляет в клетку триозофосфаты, О2 и АТФ. Интересно отметить, что фонд (пул) триозофосфатов самого хлоропласта в темноте пополняется за счет триозофосфатов гликолиза (рис.1.10).
Рис. 1.10. Метаболические реакции в хлоропластах
В цитоплазме происходит использование триозофосфатов хлоропластов и гликолиза на синтез сахарозы. На активность хлоропластов влияют также гликолитическая фосфоглицериновая, яблочная и аспарагиновая кислоты.
Неорганический фосфат поступает в хлоропласт с помощью переносчика, находящегося во внутренней мембране хлоропласта, в котором перенос фосфата в строму хлоропласта сопряжен с выходом триозофосфатов.
Митохондрии осуществляют два основных процесса: цикл Кребса и окислительное фосфорилирование. Первый из них локализован в матриксе митохондрий, а система транспорта электронов и фосфорилирования находится во внутренней мембране.
Начальное соединение цикла Кребса – пировиноградная кислота – образуется в процессе гликолиза в растворимой фазе клетки, поэтому она должна проникнуть через мембраны митохондрий в матрикс. Монокарбоновые кислоты (в т. ч. и пируват) проходят через мембраны митохондрий довольно легко с помощью переносчика. Для транспорта неорганического фосфата, ди- и трикарбоновых кислот также имеются соответствующие транспортные механизмы.
Очень распространенной формой транспорта является обмен с участием яблочной кислоты или неорганического фосфата. Транспорт адениновых нуклеотидов через внутреннюю мембрану митохондрий осуществляется также специальным переносчиком.
Поскольку многие интермедиаты цикла Кребса (пируват, малат, щавелевоуксусная кислота) могут синтезироваться в цитоплазме в процессе гликолиза и в хлоропластах, то деятельность митохондрий тесно связана с функционированием этих клеточных образований.
Митохондрии используют на собственные транспортные и синтетические процессы лишь часть синтезируемой АТФ; основное количество АТФ отдается на нужды клетке.
Необходимо отметить тесную взаимосвязь деятельности хлоропластов и митохондрий: 1) начальные продукты фотосинтеза и конечные продукты дыхания сходны; 2) конечные продукты фотосинтеза являются субстратами для дыхания; 3) в обоих процессах используются общие вещества для преобразования энергии – неорганический фосфат, пиридиннуклеотиды, аденилаты, триозофосфаты; 4) в обоих процессах фосфорилирование регулируется АДФ и неорганическим фосфатом.
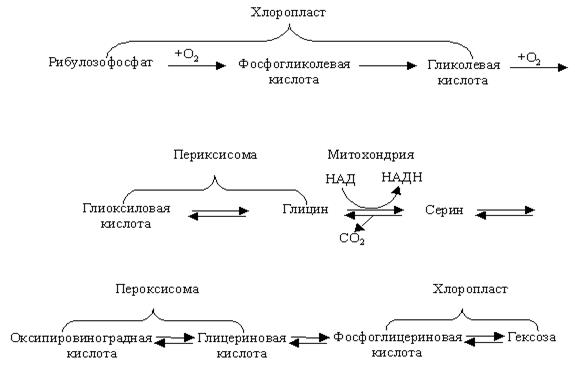
Функционирование митохондрий и хлоропластов связано с деятельностью других органелл, в частности пероксисом. Пероксисомы вместе с хлоропластами участвуют в реакциях гликолатного пути, восстановление углерода:
|
Начальные и конечные этапы цикла происходят в хлоропластах; промежуточные – в пероксисомах; процессы, связанные с синтезом АТФ через образование НАДН и выделением СО2, – в митохондриях. Кислород потребляется в двух реакциях цикла: в хлоропластах при образовании фосфогликолата и в пероксисомах при окислении гликолата в глиоксилат. Поскольку на свету эти превращения ответственны за стимуляцию поглощения О2 тканями зеленых растений, они получили название фотодыхания (см. раздел 2.14).
Метаболические системы цитоплазмы, митохондрий и хлоропластов взаимодействуют также в механизмах поддержания постоянства рН в цитоплазме. Для этого существует три механизма: буферные системы клетки, биохимический «рН-стат» и биофизический «рН-стат» (протонная помпа).
Основными буферами растительных клеток являются фосфатный и карбонатный, обнаруживающие буферные свойства при рН 5,4–6,2. В кислой области рН (3,5–5,5) проявляются буферные свойства органических кислот и их солей. Часть буферной емкости клетки определяется амино- и карбоксильными группами белков цитоплазмы (изоэлектрическая точка при рН 5,1–5,3).
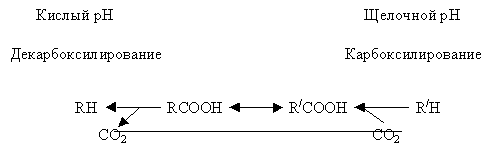
Механизм поддержания рН с помощью биохимического «рН-стата» состоит в поддержании равновесия между реакциями карбоксилирования и декарбоксилирования в цитоплазме, имеющие различные оптимумы рН, причем величины оптимальных рН могут различаться на 0,1–0,2 единицы (рис. 1.11).
|
| Рис. 1.11. Биохимический рН-стат |
Третий механизм – локализованные в мембранах системы активного транспорта ионов Н+ (или ОН–) – протонные помпы (насосы). Протонные помпы функционируют в сопрягающих мембранах митохондрий и хлоропластов (редокс- и АТФ гидролизующие Н+-помпы). Эти мембраны содержат от 2 000 до 4 000 молекул гидролитической помпы и 500–3 000 молекул редокс-протонного насоса. Протонные помпы мембран и митохондрий выносят Н+ из матрикса (подкисление цитоплазмы вблизи митохондрий) Н+-помпы хлоропластов на свету, наоборот, подщелачивают цитоплазму, поскольку направлены из цитоплазмы в строму. Плазмалемма эукариот содержит АТФ гидролизующую Н+-помпу, выкачивающий ионы Н+ из цитоплазмы в клеточную стенку. Из цитоплазмы в вакуоль направлены также Н+-помпы тонопласта.
В последние годы появились экспериментальные данные о влиянии митохондрий на фотохимическую активность хлоропластов. Исследования проводились in vitro, т. е. органоиды выделялись из растительных тканей проростков гороха (митохондрии из корней, хлоропласты из листьев); смотрели происходящие процессы при их смешивании. При внесении митохондрий в суспензию хлоропластов происходило ингибирование фотофосфорилирования в хлоропластах. Показано, что эффектор находящийся внутри клетки (вакуоль + цитоплазма), обладает протекторными свойствами только по отношению к транспорту электронов, но не способен снять обусловленное митохондриями ингибирование фотофосфорилирования.
При хранении внутриклеточной жидкости на холоде (0–4) в течение 5–6 суток или при нагревании в течение 5 минут до 40 оС активность не теряется; кипячение в течение 5 минут приводит к необратимой потере активности. Это говорит в пользу белковой природы эффектора (протектора). Этот фрагментарный пример также свидетельствует о взаимосвязанности процессов, происходящих в двух органоидах – хлоропластах и митохондриях.
Ядерно-цитоплазматические взаимодействия обеспечивают создание структуры ядра, хранение и считку генетической информации, синтез и обновление всех белков клетки, т. е. осуществляют контроль над процессами жизнедеятельности клетки. Они точно скоординированы во времени и пространстве. Все белки синтезируются на рибосомах в цитоплазме, в которую из ядра поступают мРНК, тРНК, рРНК. Из растворимой фазы клетки и локализованных в ней органоидов в ядро поставляются белки и липиды для структур ядра и ядрышка, белки субъединиц рибосом, предшественники нуклеиновых кислот, АТФ, ГТФ, ионы. В процессах транспорта важную регулирующую роль играют поровые комплексы ядерной оболочки. В цитоплазме происходит сборка рибосом и полирибосом и осуществляется активация аминокислот, необходимых для синтеза белков.
Помимо взаимодействия с растворимой фазой цитоплазмы ядро контролирует деятельность митохондрий и хлоропластов, обладающих собственными геномами, отличающимися от ядерного. В настоящее время не вызывает сомнений, что синтез мембранных белков и других ферментных систем митохондрий и хлоропластов требует кооперативной активности ядерного генома и геномов этих органоидов. Так, ядро кодирует и обеспечивает синтез в цитоплазме примерно 90 % митохондриальных белков. Структура митохондриальной ДНК определяется ею самой, но системы, от которых зависит функционирование этой ДНК – репликации и транскрипции, – контролируется ядром. Ядерный геном полностью отвечает за синтез ферментов матрикса и наружной мембраны митохондрий. Они синтезируются на цитоплазматических рибосомах. В то же время белки рибосом и внутренних мембран контролируются как ядерным, так и митохондриальным геномами. Предполагается, что белки, синтезируемые в цитоплазме, активируют работу митохондриальных рибосом, а продукты митохондриальной трансляции могут избирательно влиять на активность ядерного генома, определяющего синтез систем транскрипции и трансляции митохондрий.
Информационная емкость хлоропластного генома значительно больше митохондриального. Как и в случае с митохондриями, синтез белковых компонентов хлоропластов находится под двойным контролем ядра и хлоропластной ДНК, взаимодействие которых обуславливает синтез белков оболочки, тилакоидов и стромы, ряда компонентов I и II фотосистем и др. Особенностью деятельности генома хлоропластов является его регуляция светом, который через систему фоторецепторов хлоропласта контролирует работу белоксинтезирующей системы. Синтез белков-рецепторов (фитохром, протохлорофиллид) может контролироваться ядром. Другим каналом регуляции является регуляция деятельности хлоропластного генома на уровне транскрипции продуктами фотосинтеза. Возможно также гормональная регуляция развития хлоропластов цитокинином.
Таким образом, можно видеть тесную взаимосвязь функционирования отдельных органоидов как между собой, так и с цитоплазмой.
Предмет ГП как отрасли права.
ГП— это совокупность правовых норм, регулирующих общественные отношения имущественного и личного неимущественного характера, возникающие между участниками гражданского оборота, основанные на принципах гражданского законодательства.
ГП как отрасль права регулирует правоотношения, возникающие между гражданами и юридическими лицами. Следует иметь в виду, что участниками гражданского оборота, наряду с гражданами и юридическими лицами, выступают государство, государственные органы и органы местного самоуправления.
Как все отрасли права, гражданское право имеет свои предмет и метод правового регулирования, отличающие гражданское право от других отраслей.
Предмет гражданского права составляют общественные отношения, урегулированные нормами гражданского права. В предмет гражданского права входят две группы общественных отношений: имущественные и личные неимущественные.
Имущественные отношения представляют собой отношения участников гражданского оборота по поводу имущества или иных материальных благ, включая владение, пользование, распоряжение имуществом, а также отношения, связанные с выполнением работ или оказанием услуг. Имущественные отношения обладают экономическим содержанием и составляют следующие категории:
· вещные — отношения собственности на вещи (имущество) и отношения, возникающие в связи с законным нахождением имущества у субъектов, не выступающих его собственниками;
· обязательственные — связанные с переходом имущества от одного участника отношений к другому, возникающие из договора, вследствие причинения вреда, а также других оснований, предусмотренных гражданским законодательством.
Личные неимущественные отношения представляют собой отношения по поводу неимущественных благ. Данные отношения находятся в неразрывной связи с личностью участника гражданских правоотношений, они неотделимы и неотчуждаемы от личности (честь, достоинство, деловая репутация).
Дата добавления: 2014-12-22; просмотров: 1610;