Острое угасание условного рефлекса.
Разовое угасание при длительном действии условного раздражителя характерно отличается от многократного систематического угасания, производимого в течение многих дней без применения безусловного раздражения. В первом случае угасание проходит бесследно после отдыха, во втором же случае этого не наблюдается.
Для иллюстрации на рис. 161 приводим случай острого угасания оборонительного условного сгибания правой передней лапы при длительном применении условного раздражителя (стук метронома). У этой собаки был выработан еще другой однородный условный рефлекс на той же лапе на тов 200 щ. Из рисунка видно, что после угасания рефлекса на стук метронома тон все еще вызывает его. Это указывает на то, что угасание рефлекса в данном случае не зависело ни от утомления, ни от торможения двигательных
элементов коры, общих для этих рефлексов. Такой результат получается, если угасить условный рефлекс путем многократного повторения через короткие интервалы. Рефлекс сначала вызывается много раз, а затем, постепенно ослабевая, угасает. При этом другие однородные рефлексы не угасают. Такой пример приводится на рис. 160. У собаки было выработано несколько однородных рефлексов на правой передней лапе (на тон 200 гц, стук метронома и свет электрической лампы). После того, как угасили рефлекс на тон 200 гц, метроном и свет вызывали рефлекс. А когда был угашен рефлекс на свет, рефлекс на тон восстановился, несмотря на то, что до этого все время вызывался рефлекс на свет. Из этого опыта ясно видно, что угасание условного рефлекса зависело не от утомления или торможения двигательных элементов коры, общих для однородных рефлексов.
Можно было предположить, что данное угасание зависит от утомления начальных нервных элементов поступательных временных связей, воспринимающих условное раздражение. Мы специальными опытами доказали, что угасание не может зависеть от этого. Мы обнаружили, что длительное действие какого-либо необычного звука, вызывающего ориентировочную реакцию, не теряет своего отрицательного действия на условный оборонительный рефлекс и после того, как он перестает вызывать ориентировочную реакцию. Можно звонить или включать метроном на десятки минут и в это время условный звук, например тон 600 гц, не будет давать условного рефлекса. Отсюда вывод, что при остром угасании условного рефлекса, когда условный звук продолжается не более нескольких минут, воспринимающие данный звук корковые элементы, прежде всего сенсорные нейроны, не должны утомляться или тормозиться. Следовательно, при остром угасании утомляются или тормозятся не конечные нервные элементы поступательных временных связей, т. е. проекционные пирамиды, и не начальные пункты их — звездчатые клетки и другие воспринимающие элементы, а сами временные связи (Беритов, 6).
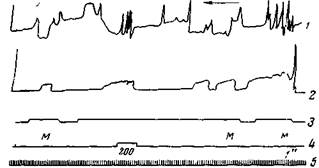
Рис. 161. Влияние угасания одного условного оборонительного рефлекса на другой однородный рефлекс
Собака Боб. 1 —движение головы; 2 — движение правой передней лапы; з — отметка раздражения — метроном; 4 — то же, тон 200 гц; 5 — время. Длительным двухминутным действием сигнала (метронома) производится угасание условного рефлекса на правой передней лапе. Приблизительно через минуту от начала этого сигнала данный рефлекс угас. В это время применяется другой условный сигнал — тон 200 гц, который вызывает значительный условный рефлекс. После второго перерыва в применении метронома всего на 12 сек. угасший метрономный рефлекс восстановился, но он очень слаб
Однако специальными опытами мы обнаружили, что когда у собаки было образовано два однородных рефлекса на одной лапе, то при угасании одного, более старого и сильного рефлекса, путем многократного повторения подряд через каждые 10—12 сек., также ослабевает другой, сравнительно слабый и молодой рефлекс. Это так называемое вторичное угасание другого рефлекса
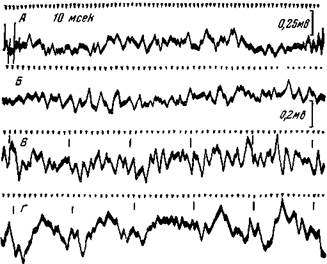
Рис. 162. Электрическая активность коркового анализатора при угасании условного рефлекса
Кошка с электродами, вживленными в слуховой области, после выработки условного пищевого рефлекса на звуковые толчки 10 в сек. А — электрическая активность слуховой области во время еды; Б — электрическая активность там же в отсутствие еды и условного сигнала; В — реакция на условные звуковые щелчки через 5 сек. от начала их действия, когда возник условный рефлекс; Г — 6-е применение условного раздражения без подкрепления (интервал между применениями звуковых толчков равнялся 1 мин.) через несколько сек. от начала его действия,^ при этом условный рефлекс не возник (Ройтбак)
проходит быстро. Его можно заметить только в том случае, если второй рефлекс испытывается во время условного раздражения угашенного рефлекса или спустя несколько секунд после него. Так как для всех этих рефлексов общими являются конечные пункты временных связей и затем весь нисходящий нервный путь, а сами временные связи различны, то, очевидно, в данном случае угнетающее действие одного рефлекса на другой происходит в этом двигательном аппарате. Но мы знаем, что даже после абсолютного угасания условного сгибания лапы, электрокожное раздражение последней вызывает безусловный рефлекс сгибания в интенсивной форме. Отсюда следует, что в данном случае утомление или торможение претерпевает конечный пункт в коре, а не подкорковые двигательные механизмы (Беритов, 6).
Итак при остром угасании условного рефлекса заметно ослабевает в коре не только деятельность временных связей, но и конечных пунктов поступательных временных связей.
Таковы были наши фактические основания для суждения о локализации угашения в корковом субстрате условного рефлекса.
Электроэнцефалографические исследования условного рефлекса, проведенные в последнее время, во многом подтвердили означенные выше теоретические предположения. Прежде всего Ройтбаком (7,8) было показано, что после угасания ориентировочного рефлекса на звуковые щелчки действие последних на кору продолжается в течение многих минут. Они все время вызывают в слуховой зоне коры первичные ответы в виде положительного потенциала и последующего альфа-подобного отрицательного потенциала. Этими наблюдениями доказано, что воспринимающая звездчатая система клеток в слуховой области в ответ на звуковое раздражение продолжает активироваться без утомления или торможения как во время ориентировочной реакции, так и долгое время после ее прекращения.
Далее, оказалось, что и при угашении условного пищевого рефлекса на звуковые щелчки, вызываемые ими, первичные ответы не ослабевают, а,
наоборот, усиливаются. Как положительный, так и отрицательный потенциалы становятся более интенсивными: ослабевают же те быстрые колебания потенциала - бета-волны, которые претерпели усиление при образовании условного рефлекса (рис. 162). Так как усиленная активация бета-волн была связана с образованием временных связей, то ослабление их при угасании должно указывать на утомление или торможение временных связей.
Наконец Ройтбак показал, что при длительном действии условного раздражения, на которое угас пищевой рефлекс, животное переходило в сонное -состояние, и в это время медленные отрицательные потенциалы возникали и в других областях коры. Одновременно уменьшались амплитуда и ритм бетаволн (рис. 163). Следовательно, при угасании пищевого условного рефлекса мы имеем дело с общим торможением нервных кругов, производящих эти бета-волны. А так как медленные потенциалы возникают при активации пирамидных дендритов, то, надо полагать, согласно дендритной гипотезе торможения что общее торможение быстрых потенциалов происходит вследствие электротонического распространения возникающих при этом дендритных токов до тела пирамидных клеток и их анэлектротонического действия на прилегающие к ним синапсы.
Так как при угасании пищевого ориентировочного условного рефлекса на звук медленные потенциалы возникают не только в слуховой области, куда приходят периферические импульсы по специфической системе, но и по всей коре, то нельзя допустить, что активация всей дендритной массы происходит через коллатерали нервных кругов условного рефлекса.
Очевидно, через коллатерали временных связей угнетаются лишь нервные связи двигательного анализатора с проекционными пирамидами, которые вызывают периферическую реакцию в рабочем органе. Это угнетение, по-ви-
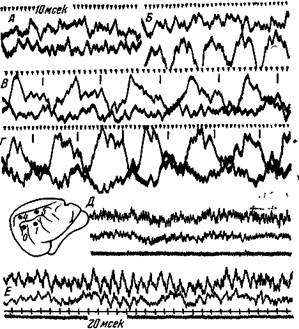
Рис. 163. Электрическая активность коркового анализатора после угасания пищевого ориентировочного рефлекса на условные звуковые толчки 10 в сек. Та же кошка. Потенциалы отводятся от пунктов 1 и о (верхние кривые) и от пунктов 4 и б (нижние кривые) А—«спонтанная» активность, кошка не спала; Б — кошка стала дремать; В—кошка спокойно сидела, через 1 сек. от начала действия звуковых толчков; Г — через несколько сек. действия звуковых толчков; Д и Е — другая кошка: Д — до раздражения, Е — через 25 сек. от начала действия условных звуковых толчков, когда они перестали вызывать пищевой ориентировочный рефлекс (Ройтбак)
димому, происходит вследствие активации базальных дендритов проекционных пирамид. Активация должна была заметно усилиться вследствие развития обратных связей. На электроэнцефалограмме это проявляется в виде усиления положительных потенциалов. Ассоциационные нейроны обратных связей лежат в двигательном анализаторе — конечном пункте поступательных временных связей, и поэтому возбуждение этих нейронов должно активировать через их коллатерали прежде всего находящиеся в соседстве базальные дендриты проекционных пирамидных нейронов.
Что же касается возникновения медленных потенциалов и угнетения быстрых вне слуховой области по всей коре, то это должно происходить, как считает и Ройтбак, путем вовлечения тормозящего диэнцефалического механизма. Импульсы из этого механизма по неспецифической системе достигают верхушечных дендритов пирамидных нейронов всей коры и вызывают в них местное возбуждение, производящее при отведении с поверхности коры медленные отрицательные потенциалы. За это говорит, между прочим, то обстоятельство, что медленные отрицательные потенциалы в коре протекают по типу альфа-волн, которые по своему происхождению связаны с деятельностью кортико-таламо-кортикальных путей.
Активация диэнцефалического тормозящего механизма, очевидно, обусловливается центрифугальной импульсацией из коры по экстрапирамидным путям. Так можно думать прежде всего потому, что у кошек без неокортекса ориентировочная реакция на звуки полностью не угасает и после их многократного применения, в то время как у нормальных кошек она угасает после нескольких повторений звука. Следовательно, тормозящий диэнцефалический механизм в отношении ориентировочной реакции не активируется непосредственно через восходящие пути. Двигательные центры ориентировочной реакции головы и глаз лежат в среднем мозге. При периферических раздражениях они активируются в первую очередь. И если на нормальных животных ориентировочная реакция угасает очень быстро, а у животных без неокортекса или вообще без коры она не угасает, то это лучше всего свидетельствует о том, что деятельность этих двигательных механизмов регулируется из коры. Вероятно, при угасании эта регуляция осуществляется через диэнцефалический тормозящий механизм, который, по-видимому, тормозит кору одновременно с торможением двигательных механизмов ствола мозга.
Дата добавления: 2017-01-13; просмотров: 1166;