Экологические группы по отношению к свету
На Земле существует огромное разнообразие местообитаний, отличающихся по световому режиму (интенсивность света, его спектральный состав, продолжительность освещения, распределение света в пространстве и времени). Растения живут и при очень сильном освещении в пустынях и высокогорьях, и в темных пещерах и подводных глубинах. В процессе длительной эволюции растительные организмы приобрели эффективные приспособления к успешной жизни при световом режиме их местообитаний.
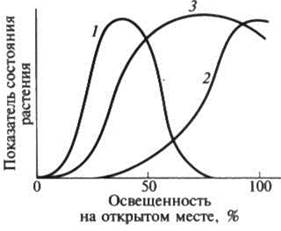
В. Н. Любименко предложил выделять три группы растений по отношению к свету: гелиофиты(от греч. helios — солнце), или световые (светолюбивые), сциофиты(от греч. skia — тень), или теневые (тенелюбивые), и теневыносливые. Они отличаются, прежде всего, положением светового оптимума и кардинальных точек светового довольствия (рисунок 1). У гелиофитов оптимальное световое довольствие находится в области почти полного освещения и сильное затенение их угнетает. Световое довольствие этих растений близко к 100 %, они растут на открытых местах, где инсоляция велика и растительный покров изрежен. К этой группе относятся виды жарких пустынь, тундр, высокогорий, скал и каменистых осыпей, рудералы пустырей и обочин дорог, большинство культурных растений открытого грунта и сорняков, многие прибрежные и водные растения с
|
Рисунок 1 - Экологические оптимумы растений по отношению к свету (по
Т. К. Горышиной, 1979):
1 — тенелюбивый вид; 2 — светолюбивый вид; 3 — теневыносливый вид
надводными листьями, деревья первого яруса и ранневесенние травы листопадных лесов. Гелиофитами являются, в частности, сосна обыкновенная (Pinus sylvestris), береза повислая (Betula pendula), мать-и-мачеха (Tussilagofarfara), очиток едкий (Sedum acre) и др.
Однако границы между дискретными экологическими группами условны: гелиофиты образуют непрерывный ряд по изменению светолюбия. Действительно, некоторые из них могут расти в слабозатененных условиях. Поэтому различают облигатные гелиофиты, не выносящие затенения, и факультативные, хорошо растущие и при небольшом затенении. Кроме того, их экологические предпочтения пластичны в онтогенезе и конкурентной среде. С хозяйственной точки зрения, поскольку полевые сорняки большей частью светолюбивы, действенное средство борьбы с ними — затенение. Засорены же в первую очередь поля с зерновыми, дающими мало тени.
Теневыносливые растения имеют широкую экологическую амплитуду по отношению к свету, и их точнее называть свето- и теневыносливыми. Максимальное световое довольствие (Lmax) их равно 100%, а минимальное (Lmin) разнообразно и характеризует способность углубления в тень. Как правило, световая кривая их асимметрична: они лучше растут при большой освещенности, но хорошо адаптируются и к слабому свету. Экспериментально показано, что многие растения затененных мест при предоставлении им несколько большего освещения развиваются лучше, т.е. они именно теневыносливы. Это распространенная и очень пластичная группа. Сюда относятся, например, многие лесные травы. А в соответствии с континуальным изменением данного признака у лесоводов давно существуют шкалы теневыносливости древесных пород.
Сциофиты имеют оптимум в области слабой освещенности и не выносят сильного света. Он подавляет их, так как интенсивно разрушает хлорофилл, а они медленно его производят. Кроме того, теневые растения при сильной инсоляции так быстро теряют влагу при транспирации, что вынуждены закрывать устьица, что приводит к торможению фотосинтеза и голоданию. Поэтому сциофиты не растут на полном свету (у них всегда Lmax < 100 %). Также они заходят в более глубокую тень, чем теневыносливые виды, и их минимальное световое довольствие всегда ниже. Сциофиты предпочитают ослабленное освещение. К тенелюбивым видам принадлежат растения сильно затененных мест. При этом исходно затененные местообитания (пещеры, расщелины скал, водные глубины и т.п.) сравнительно редки. В основном в экосистемах затенение — результат перехвата света более высокими растениями. Поэтому среди сциофитов много лесных видов нижних ярусов (мхи, папоротники, ряд трав, например грушанки — Pyrola, и др.). Оказавшись на вырубке, эти растения желтеют из-за разрушения хлорофилла и вскоре погибают. К тенелюбам относятся также многие комнатные растения, являющиеся выходцами из травяного покрова или эпифитов тропических лесов. По способности расти на свету, среди сциофитов тоже можно выделить факультативные и облигатные виды.
Степень теневыносливости или светолюбия не является строгим видовым признаком. Внутривидовые анатомо-морфологические и физиологические различия выражены у особей, растущих в разных условиях освещенности, — в глубине леса и на опушке, в густом травостое и при одиночном произрастании, на северной и южной стороне стволов и т.д. Способность растений выносить недостаток света в большой степени зависит от комплекса других экологических факторов. В оптимальных климатических и почвенных условиях теневыносливость вида обычно выше. Этим объясняются закономерные географические изменения по отношению к свету широко распространенных видов. Например, в северных частях ареала виды средней полосы более светолюбивы, чем в южных.
Световое довольствие увеличивается и при подъеме в горы, что отмечено на примере дикого ячменя (Hordeum): на высоте 180 м он выносит 1/9 полного освещения, а на 2500 м ему требуется не менее 1/5 (Т. К. Горышина, 1979). Возрастание светолюбия при понижении температуры по мере продвижения растений к полюсу — одна из причин смены местообитаний некоторых видов на более осветленные. Так, лесная черника (Vaccinium myrtillus) в тундре хорошо растет на открытых местообитаниях, а сныть (Aegopodium podagrariа), один из наиболее теневыносливых видов дубрав, на севере России обитает на опушках и лугах. Аналогично отношение растений к свету и при подъеме в горы. Особо меняется светолюбие у растений арктической зоны, где летом освещение круглосуточно: в холодные ночи они лучше используют свет слабой интенсивности (как тенелюбы), а днем хорошо используют интенсивный свет (как светолюбы).
Формообразование растений под действием света.Растения и их части, развивающиеся на полном свету, анатомо-морфологически отличаются от теневых. В связи с этим выделяют черты гелиоморфоза (или фотоморфоза) — формообразования под действием света. Некоторые из них — видовые, другие развиваются лишь в определенных условиях. Часто эти черты сопряжены с ксероморфными признаками, что связано с напряженным водоснабжением в сильно освещаемых местах. У теневых же листьев обычно проявляются мезо- и гигроморфные признаки.
В целом побеги гелиофитов обычно более толстые, с хорошо развитой ксилемой и механической тканью. Корни их более длинные и разветвленные, а у бобовых на них развивается больше клубеньков и они крупнее. Большинство светолюбивых древесных пород — анемохоры с мелкими семенами, и всходы их лучше выживают на оголенной почве или под изреженным покровом низких трав. В ажурных кронах этих деревьев свет ослабевает постепенно, а сильное развитие осевого побега у некоторых видов приводит к пирамидальноcти кроны. В сомкнутом сообществе нижние ветви этих деревьев рано отмирают, стволы очищаются и кроны располагаются высоко. Для гелиофитов характерны также выраженный фототропизм и возникновение асимметричности при одностороннем освещении.
Прямой яркий свет тормозит рост в длину. Междоузлия трав-светолюбов нередко укорочены, что ведет к образованию розеточности. А сочетание укорачивания междоузлий с сильным ветвлением приводит к подушковидности. В связи с медленным ростом растения-подушки по долговечности сравнимы с деревьями.
При недостатке света у многих растений (особенно у гелиофитов с запасающими органами) стебли аномально вытягиваются, приобретают бледную окраску и мелкие листья. Утолщение и одревеснение их в затенении отстает от нарастания в длину. Развившиеся в темноте побеги вытянутые, бледные и не способные поддерживать себя в вертикальном положении, называют этиолированными (по франц.—«хилые»). Элементы этиолированности часто встречаются у растений при совместном произрастании в лесу, на лугах, в загущенном посеве. Так, у клевера (Trifolium) в густом травостое междоузлия удлинены и листья уже по сравнению с экземплярами, выросшими при меньшей сомкнутости. Многие теневые растения закрепили некоторую этиолированность в своей жизненной форме. Они нередко становятся стелющимися или ползучими, а некоторые имеют полегающие стебли, легко укореняющиеся при достаточной влажности. Если при этом сильное вытягивание стеблей может сочетаться с недоразвитием листьев, образуются столоны.
Особенности листьев световыхи теневых растений.На расположение, форму и строение листьев влияет множество факторов, но решающее значение в связи со специализацией листа как приемника света и поверхности транспирации имеют световой режим и водообеспеченность.
Размер листьев — экологически очень важный параметр. Действительно, одно из наиболее наглядных различий внешнего облика растений в разных световых условиях — неодинаковая величина их листовых пластинок. От этого в большой степени зависит количество радиации, получаемое растением. Этот признак во многом наследственно детерминирован, но также связан и с модификациями в разных условиях светообеспеченности (и обеспеченности водой).
Уже простые модели, основанные на оптимизации использования воды листом (количество граммов СО2, ассимилированного на 1 г израсходованной воды), предсказывают существование нескольких размерных классов листьев. Например, крупных у растений затененных влажных теплых мест и мелких у растении холодных или более теплых и одновременно солнечных местообитаний. При одинаковом водообеспечении у светолюбивых растений листья обычно мельче, чем у тенелюбивых (рисунок 2).
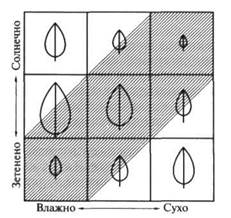
Рисунок 2 – Схема распределения размеров листьев в зависимости
от условий обитания растений (по Э. Пианка, 1981). Заштрихованный участок соответствует преобладающим в природе условиям
Это хорошо видно при сравнении родственных видов, живущих при разном освещении. Так, мелколистные колокольчики (поникший Campanula patula и круглолистный С. rotundifolia) растут на лугу, а колокольчики с широкими листьями (крапиволистный С. trachelium и широколистный С. latifolia) обитают в лесу. Лесные злаки и осоки тоже обычно представлены видами с более широкой листовой пластинкой, чем луговые. Однако все же затенение по-разному может влиять на размеры листа. У светолюбивых видов любое затенение ведет к редукции площади листьев и максимальных размеров они достигают только при самом сильном освещении (например, у подсолнечника — Helianthus annuus). Листья же теневыносливых видов, как это было показано на недотроге (Impatiens noli-tangere), при умеренном затенении становятся крупнее, но при сильном затенении мельчают. Увеличение листовых пластинок при затенении до некоторой степени компенсирует снижение интенсивности фотосинтеза ростом фотосинтезирующей поверхности. При этом еще уменьшается относительная масса не ассимилирующих органов и соответственно падает расход органики на дыхание. Однако чрезмерное увеличение площади листьев вредно, поскольку нарушает оптимальное соотношение между поверхностями листьев и корней, что может поставить растение под угрозу при ухудшении условий среды (дефиците водоснабжения, усилении инсоляции и т.д.). Морфологической адаптацией является и разное расположение листовых пластинок в пространстве в условиях избытка или недостатка света. У гелиофитов листья обычно ориентированы так, чтобы избежать избыточного полуденного света. Их листовые пластинки расположены вертикально или под большим углом к горизонту, поэтому днем листья получают лишь скользящие лучи. Среди древесных пород такое положение листьев очень характерно для эвкалиптов (Eucalyptus). Пластинки листочков сложных листьев мимоз (Mimosa) и акаций (Acacia) тоже демонстрируют такое расположение. Среди трав свойство поворачивать листья сообразно падающим лучам имеют многие степные виды.
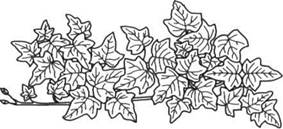
Рисунок 3 – Листовая мозаика у плюща (Hedera)
Другая адаптация, связанная с необходимостью ослабления получаемой радиации, встречается у компасных растений. Их листовые пластинки расположены в одной плоскости, которая ориентирована с севера на юг, поэтому в полдень приход радиации к листу минимален. Типичное компасное растение — дикий латук (Lactuca serriola), растущий в разных открытых местообитаниях. На сильно освещенных местах тенденцию к компасному расположению листьев приобретает и ряд обычных луговых трав (пижма обыкновенная — Tanacetum vulgare, тысячелистник обыкновенный — Achillea millefolium и др.). Однако в тени компасность теряется и ориентация листьев становится беспорядочной.
У теневыносливых растений, растущих при ослабленном освещении, напротив, листья направлены так, чтобы максимизировать поглощение света. Поэтому листья растений нижних ярусов леса часто ориентированы по направлению к просветам в пологе древостоя. Если же просветов в густом пологе нет, то листья трав и подроста расположены почти горизонтально, улавливая рассеянный свет. То же отмечается в густых луговых травостоях и нижних частях плотных крон. Известным приспособлением, уменьшающим затенение, является листовая мозаика, получающаяся при расположении листьев в одной плоскости (рисунок 3). Она характерна для подроста и нижних ветвей многих широколиственных деревьев (клен — Acer, вяз — Ulmus, липа — Тiliа). Подобное расположение листьев встречается и у трав сильно затененных лесов.
Некоторые теневые растения способны менять положение листовых пластинок при попадании яркого света. Так, избегая светового повреждения, оказавшаяся на вырубке кислица (Oxalis acetosella) складывает листья так, что их доли приобретают вертикальное положение.

Рисунок 4 – Кислица обыкновенная (Oxalis acetosella)
Многие гелиофиты имеют белесоватую окраску, что предохраняет их от чрезмерного светового и теплового воздействия. При этом поверхность органов часто имеет особенности, способствующие отражению лучей. Она может быть блестящей, покрытой восковым налетом, опушенной светлыми волосками. Характерно, что у дорсовентральных листьев отражение от нижней стороны почти в 3 раза выше, чем от верхней. Это защищает нежную губчатую паренхиму и устьица от перегрева в случае поворачивания листа ветром. Сильно ослабляют свет густое опушение, толстая кутикула и утолщенные наружные стенки клеток эпидермиса. Так, эпидермис световых растений пропускает не более 15% падающего света, а эпидермис теневых — до 98 %. В эпидермисе некоторых гелиофитов, растущих на меловых отложениях, обнаруживаются кристаллические включения, экранирующие световые лучи. То же иногда отмечается у липы (Tiliа) при избытке освещения. Защитную роль играет и гиподерма хвои сосны (Pinus) и ели (Picea).
Внутреннее строение листа гелиофитов отличается рядом черт «световой структуры» ассимиляционной ткани. Это, прежде всего, хорошо развитый столбчатый мезофилл, составляющий около 60 % от общей поверхности фотосинтезирующей ткани. Он состоит из узких длинных клеток, лежащих иногда в 2—3 ряда. Вдоль них по межклетникам свет эффективно проникает в глубь листа, достигая хлоропластов, постенно расположенных в узких клетках. Многослойная столбчатая паренхима световых листьев позволяет гелиофитам поглощать большие порции света. Многим гелиофитам свойственны также изопалисадные листья, в которых столбчатая ткань хорошо развита на обеих сторонах. Это явление часто наблюдается у растений с вертикальным положением листьев и видов на меловых выходах, отражающих падающий на них свет.
В связи с развитием мощной столбчатой ткани возрастает общая толщина листа, поэтому световые листья часто толще теневых. Обычно единица площади световых листьев имеет значительно большую массу, чем теневых, т.е. у них меньше удельная поверхность листа (площадь, приходящаяся на единицу массы). В более толстом листе гелиофитов гуще и сеть жилок, а клетки обычно расположены компактно, без крупных межклетников. Для листьев гелиофитов типична и мелкоклеточность, также обусловливающая меньшие размеры их листьев. Устьица их обычно более мелкие и многочисленные.
В целом структура светового листа обеспечивает и достаточный перехват солнечной радиации, и хорошо защищает лист от чрезмерного облучения.
У растений затененных местообитаний листья имеют черты «теневой» структуры. Листовые пластинки их более тонкие, с большей удельной поверхностью, что способствует проникновению слабого света в глубь листа. Клетки теневых листьев, в том числе устьичные, обычно довольно крупные. Устьица редкие, рассеянные и в отличие от световых листьев расположены только на нижней стороне листа. Для теневых листьев характерны также извилистые очертания клеток эпидермиса. Мезофилл теневых листьев или совсем не дифференцирован на столбчатую и губчатую паренхиму, или клетки верхнего слоя лишь несколько отличаются от нижних (не бывают узкими и длинными). А губчатая паренхима в теневых листьях всегда хорошо выражена. В ней много межклетников, что обеспечивает полное поглощение света из-за многократного рассеивания его здесь.
На уровне хлоропластов световые и теневые листья тоже имеют различия. Листья гелиофитов мелкоклеточные и потому содержат хлоропластов относительно немного. Однако на единицу объема клетки у них приходится хлоропластов больше, чем у теневыносливых видов с их более крупными клетками, хлоропластами и межклетниками. Кроме того, у световых листьев существенно большая толщина мезофилла. В итоге в условиях сильной освещенности число хлоропластов, приходящееся на единицу площади листа, оказывается в несколько раз больше, чем у растений в затенении. Так, очень много хлоропластов на 1 см2 листьев у ксерофитов аридных пустынь, обитающих в условиях высокой интенсивности света: у астрагала хивинского (Astragalus chiwensis) — 89 млн., у осоки вздутой (Carex physodes) — 72 млн.. При этом в густо наполненном хлоропластами световом листе не все пластиды получают сильный свет из-за взаимного затенения. Сами хлоропласты у гелиофитов более мелкие и светлые (с меньшим содержанием хлорофилла), а у сциофитов — крупные и темные. Так, содержание хлорофилла в одном хлоропласте теневыносливых растений в 5—10 раз больше. То же характерно для низших растений.
Физиолого-биохимическне адаптации растений к световым условиям.Основные физиологические адаптации к свету касаются фотосинтеза. Изменение концентрации хлорофилла в листьях связано с приспособлением для улавливания света. При оптимальном содержании хлорофилла световые растения создают больше сухого вещества, чем теневые. При изменении освещенности у гелиофитов количество пигментов нередко падает, причем как при увеличении, так и при уменьшении интенсивности света. В ясные солнечные дни у световых растений содержание хлорофилла снижается, и минимальное его количество отмечается при наиболее сильном дневном освещении.
Классические работы по изучению физиологических адаптации растений к условиям освещенности проделал В.Н.Любименко еще в начале XX в. Изучив более 600 видов растений, он пришел к выводу, что световые листья лучше работают при более сильном свете, а теневые — при слабом. Причем активный фотосинтез тенелюбов при слабом освещении связан с более высокой концентрацией хлорофилла (у них более темная окраска в тени). Это различие хорошо заметно при сравнении содержания хлорофилла в единице массы листа. Так, у светолюбов листья содержат 1,5 — 3 мг хлорофилла в 1 г листа, а у теневых видов — 4—6 мг (до 7 — 8 мг). В. Н. Любименко также показал, что если светолюбивые растения реагируют на затенение снижением содержания хлорофилла, то для теневыносливых типично возрастание его концентрации.
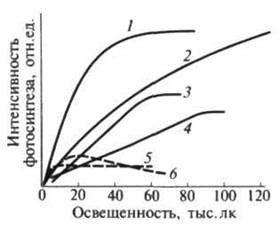
|
Рисунок 5 – Световые кривые фотосинтеза растений: 1 – калужница арктическая (Caltha arctia) (тундра); терескен серый (Eurotia ceratoides) (Памир); 3 – пролеска сибирская (Scilla sibirica); 4 – типчак (Festuca sulcata) (степь); 5 – кислица (Oxalis acetosella); 6 – сныть (Aegopodium podagraria) (дубрава)
При увеличении света фотосинтез у гелиофитов нарастает медленно (угол наклона кривой невелик), и выход кривой на плато происходит при высокой освещенности, составляющей от 1/3 до 1/2 и более от полной (рисунок 5). А очень светолюбивые виды совсем не имеют «плато насыщения», и фотосинтез у них возрастает с увеличением освещенности до максимально возможных величин. Угол наклона световой кривой у сциофитов более крутой, что свидетельствует о более эффективном использовании доступного им слабого света. У теневыносливых растений кривая выходит на плато насыщения при низкой освещенности, но максимальная интенсивность фотосинтеза при насыщающей освещенности ниже, чем у светолюбивых видов.
Дата добавления: 2015-09-21; просмотров: 12347;