Синтез и выделение нейромедиаторов, их химическая природа
Выделение нейромедиаторов пресинаптическими окончаниями нейронов напоминает секрецию эндокринных желёз, выделяющих в кровь свои гормоны. Но гормоны обычно действуют на клетки, находящиеся на удалении от самой железы, тогда как мишенями для медиаторов являются лишь постсинаптические нейроны. Поэтому у любого медиатора очень короткий путь до цели, а его действие оказывается быстрым и точным. Точности способствует наличие активных зон – специализированных областей пресинаптической мембраны, где обычно происходит выделение медиатора. Если же медиатор выделяется через неспецифические участки мембраны, то точность его действия уменьшается, а само действие замедляется. Такая картина наблюдается, например, в синапсах, образованных между нейронами вегетативной нервной системы и гладкими мышцами.
Но иногда действие медиатора не ограничено только соседней клеткой, и в таких случаях он действует как модулятор с достаточно широким спектром деятельности. А отдельные нейроны выделяют свой продукт в кровь, и тогда он действует уже как нейрогормон. Несмотря на то, что по своей химической природе многие нейромедиаторы существенно отличаются, результат их влияния на постсинаптическую клетку (т.е. возбуждение или торможение) определяется не химической структурой, а типом ионных каналов, которыми медиатор управляет с помощью постсинаптических рецепторов.
По химической структуре различают низкомолекулярные и пептидные медиаторы.
К низкомолекулярным медиаторам относятся ацетилхолин, биогенные амины, гистамин, аминокислоты и их производные. В списке медиаторов белковой природы значится свыше 50 коротких пептидов. Нейроны, выделяющие определённый медиатор, а также синапсы, в которых он используется и постсинаптические рецепторы для него принято называть …-эргическими, где вместо многоточия ставится название конкретного медиатора: например, ГАМК-эргические нейроны, адренэргические синапсы, холинорецепторы, пептидэргические структуры и т.п.
Вещества, оказывающие на постсинаптические рецепторы такое же действие, как и сам медиатор, называют агонистами, а вещества, связывающиеся с постсинаптическими рецепторами и блокирующие их без присущего медиатору действия, – антагонистами.
Для каждого медиатора существуют свои механизмы синтеза и исходные вещества. Ферменты для синтеза низкомолекулярных медиаторов находятся, как правило, в цитоплазме, а синтез происходит на свободных полирибосомах. Образовавшиеся молекулы медиатора упаковываются в синаптические пузырьки и медленным аксоплазматическим транспортом доставляются в окончание аксона. Но и в самом окончании может происходить синтез низкомолекулярных медиаторов.
 Пептидные медиаторы образуются только в теле клетки из молекул белка-предшественника при участии специальных ферментов. Их синтез происходит в эндоплазматическом ретикулуме, дальнейшие преобразования – в аппарате Гольджи. Оттуда молекулы медиатора в секреторных пузырьках попадают в нервное окончание с помощью быстрого аксонального транспорта.
Пептидные медиаторы образуются только в теле клетки из молекул белка-предшественника при участии специальных ферментов. Их синтез происходит в эндоплазматическом ретикулуме, дальнейшие преобразования – в аппарате Гольджи. Оттуда молекулы медиатора в секреторных пузырьках попадают в нервное окончание с помощью быстрого аксонального транспорта.
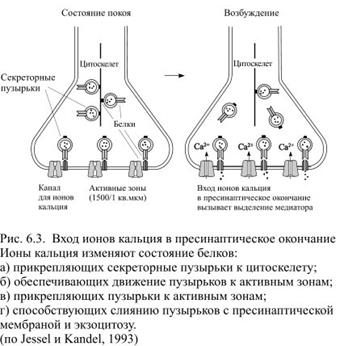
Для того чтобы молекулы медиатора попали в синаптическую щель, синаптический пузырёк должен сначала слиться с пресинаптической мембраной в её активной зоне. После этого в пресинаптической мембране образуется увеличивающееся в диаметре отверстие, через которое всё содержимое пузырька опорожняется в щель. Этот процесс называется экзоцитозом. Когда необходимости в выделении медиатора нет, большая часть синаптических пузырьков бывает прикреплена к цитоскелету.
Когда нейрон возбуждается и потенциал действия достигает пресинаптического окончания, в нём открываются потенциалзависимые каналы для ионов кальция. Роль ионов кальция состоит в том, чтобы преобразовать вызванную возбуждением нейрона деполяризацию в неэлектрическую активность – выделение медиатора. Без входящего тока ионов кальция нейрон фактически лишается своей выходной активности. Кальций нужен для взаимодействия белков мембраны синаптических пузырьков с белками плазматической мембраны аксона. В результате взаимодействия этих белков синаптические пузырьки перемещаются к активным зонам и прикрепляются к плазматической мембране. Только после этого начинается экзоцитоз (рис. 6.3).
Небольшое количество медиатора выделяется и без возбуждения нейрона, происходит это малыми порциями – квантами. В большинстве синапсов ЦНС после вхождения ионов кальция в пресинаптическое окончание выделяется от 1 до 10 квантов медиатора, поэтому одиночные потенциалы действия практически всегда оказываются подпороговыми. Количество выделяемого медиатора увеличивается, когда к пресинаптическому окончанию поступает серия высокочастотных потенциалов действия. В этом случае растёт и амплитуда постсинаптического потенциала, т.е. происходит временная суммация.
После высокочастотной стимуляции пресинаптического окончания наблюдается повышение эффективности синаптической передачи в течение нескольких минут, а у отдельных нейронов ещё дольше – до часа, когда в ответ на одиночный потенциал действия медитора выделяется больше, чем обычно. Это явление получило название посттетанической потенциации. Объясняется оно тем, что при высокочастотной или тетанической стимуляции растёт концентрация свободного кальция в нервном окончании, что вызывает повышенное отхождение синаптических пузырьков от цитоскелета и усиление экзоцитоза.
Повышение эффективности синаптической передачи является одним из механизмов образования памяти, а накопление ионов кальция в пресинаптическом окончании можно рассматривать как один из способов хранения информации о предшествующей высокой активности нейрона.
Дата добавления: 2015-01-21; просмотров: 5966;