Энергетика. Синтез АТФ в митохондриях. Молекулярные механизмы процессов энергетического сопряжения.
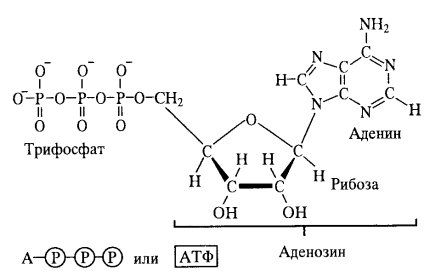
Как известно, живые системы функционируют благодаря использованию различных видов энергии трансформированием их в энергию химических связей. В клетке химическая энергия запасается в виде так называемых «высокоэнергетических» метаболитов. Наиболее важным таким метаболитом, макроэргом, обеспечивающим энергией большинство энергозависимых процессов в клетке, является нуклеотидный коэфермент аденозинтрифосфат АТФ ( рис.1). В молекуле АТФ цепочка из трех фосфатных остатков (α, β, γ) связана 5΄-ОН-группой аденозин. Рибоза связана с α-фосфатом устойчивой фосфорноэфирной связью. Три фососфатных остатка связаны между собой менее устойчивыми фосфоангидридными связями. При физиологических значениях рН АТФ несет четыре отрицательных заряда. Изменение свободной энергии ΔG0 гидролиза фосфоангидридных связей составляет 30-35 кДж/моль. В клетке, в физиологических условиях измение свободной энергии гидролиза АТФ еще выше и может доходить до 50 кДж/моль. Гидролиз АТФ в клетках сопровождается выделением свободной энергии, которая расходуется на выполнение эндергонических (энергозатратных) процессов, таких как биосинтез, движение, транспорт и др. Соответственно, синтез АТФ является высоко эндергонической реакцией, поэтому он должен сопрягаться высоко экзергоническим процессом. Молекула АТФ является самым важным и универсальным энергетическим посредником, которая обеспечивает генерирование химической энергии и ее использование для выполнения биологических функций у всех организмов. Остальные нуклеозидтрифосфатные коэферменты, химически похожие на АТФ ( ГТФ, ЦТФ, УТФ) выполняют в метаболических процессах другие функции.
|
Рис.1. Структура молекулы АТФ (кольман,125)
В ходе эволюции сформировались два важных способа синтеза АТФ, которые реализуются в живых клетках. Наиболее эффективный способ синтеза АТФ – это использование энергии градиента электрохимического потенциала биологических мембран для образования АТФ из АДФ и неорганического фосфата. Энергия для создания такого градиента возникает в результате окислительно-восстановительного процесса в результате окисления химических субстратов (окислительное фосфорилирование) или под действием световой энергии ( фотофосфорилирование).
Второй, эволюционно более ранний способ синтеза АТФ осуществляется в анаэробных условиях. В этом случае перенос фосфатной группы на АТФ осуществляется через метаболит с высоким потенциалом переноса фосфатных групп. В качестве примера можно привести реакцию синтеза АТФ из креатинфосфата в мышечных клетках.
По способу преобразования энергии организмы (клетки) подразделяются на большие 2 группы : автотрофы и гетеротрофы. Автотрофы для синтеза АТФ используют энергию электромагнитных (световых) волн или . Гетеротрофы – для синтеза АТФ используют энергию химических связей органических субстратов.
Синтез АТФ у автотрофных организмов мы подробно будем говорить на лекции, посвященному фотосинтезу. Сейчас приступим к рассмотрению процесса синтеза АТФ у аэробных организмов - окислительного фосфорилирования. Впервые синтез АТФ, сопряженный с внутриклеточным дыханием был обнаружен в начале 30-ых годов 20 века российским биохимиком В. А. Энгельгардом. Другой россиянин, В.А. Белицер, в 1939 году предположил, что окислительное фосфорилирование АДФ сопряжено с переносом электронов в дыхательной цепи. Американские ученые А.Ленинджер и Е. Кеннеди в 1949 году показали, что процесс окислительного фосфорилирования протекает в митохондриях.
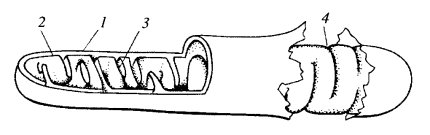
Из курса цитологии вам известно, что митохондрии – это органеллы клетки палочкообразной или округлой формы, сравнительно больших размеров ( до 2-3 мкм). Митохондрия окружена двойной элементарной мембраной, толщина каждой мембраны около 7-8 нм. Митохондрия имеет два внутренних отсека (компартмента): пространство между внутренней и внешней мембраной (межмембранное пространство ) и матрикс, пространство, ограниченное внутренней мембраной. Внутренняя мембрана образует многочисленные складки и выступы (кристы), за счет чего достигается увеличение поверхности внутренней мембраны (рис.2). В митохондриях за счет окислительной деградации питательных веществ синтезируется большая часть необходимого клетке АТФ.
|
| Рис. 2. Схема общей организации митохондрии 1 — внешняя мембрана; 2 — внутренняя мембрана; 3 — впячивания внутренней мембраны - кристы; 4 — места впячиваний, вид с поверхности внутренней мембраны |
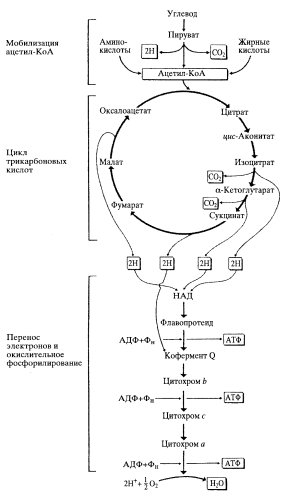
Главной функцией митохондрий является захват богатых энергией субстратов (жирные кислоты, пируват, углеродный скелет аминокислот) из цитоплазмы и их окислительное расщепление до СО2 и Н2О, сопряженнное с синтезом АТФ. Соответственно, в митохондриях локализованы следующие метаболические процессы: 1) превращение топливных субстратов в ацетил-КоА 2) восстановление НАДН и ФАДН2 в цикле Кребса 4) синтез АТФ в дыхательной цепи ( рис. 3). Реакции двух первых процессов локализованы в матриксе, а дыхательная цепь располагается на внутренней мембране митохондрий. На этой мембране митохондрии находятся молекулярные переносчики электронов и ферменты, составляющие дыхательную (электронно-транспортную) цепь. Компоненты дыхательной цепи осуществляют перенос электронов от НАДН или восстановленной формы убихинона QН2 на молекулярный кислород. Вследствие большой разницы окислительно-восстановительных потенциалов (ОВП) доноров (НАДН2 и QН2 ) и акцептора (О2), реакция переноса электронов является высоко экзергонической. Большая часть освобождающейся энергии расходуется на создание градиента концентрации протонов, которую фермент АТФ-синтаза использует для образования молекулы АТФ.
|
| Рис. 2. Схематичное изображение метаболических процессов в митохондриях |
Основные компоненты ЭТС митохондрий следующие: комплекс 1 (НАДН: убихинон –оксиредуктаза или НАДН-дегидрогеназа); комлекс 11 (сукцинат : убихинон-оксиредуктаза или сукцинатдегидрогеназа); комплекс 111 (убихинол : цитохром с – оксиредуктаза); комплекс 1У (цитохром с : кислородоксиредуктаза); цитохром с; АТФ- синтетаза; АДФ – АТФ – транслоказа; убихинон..
Перечисленные компоненты ЭТС на мембране митохондрий располагаются в следующих стехиометрических соотношениях: комплекс 1: комплекс 2 : комплекс 3: комплекс 4 как 1 : 2 : 3 : 6. На каждый комплекс 1У приходится по 1 молекуле АТФ-азы и по 3-5 молекул АДФ-АТФ – транслоказы.
Перенос электронов по дыхательной цепи. Электроны, окисляющие кислород, проходят, по меньшей мере, десять промежуточных окислительно-восстановительных систем, большинство из которых представлены простетическими группами в комплексах 1, 111, 1У. Молекулярные переносчики (НАДН, флавопротеины, содержащие ФАД и ФМН в качестве простетических групп, убихинон) акцептируют и отдают по 2 электрона. Негемовые железосерные белки и цитохромы, содержащие порфириновые простетические группы, переносят по одному электрону ( рис. 3).
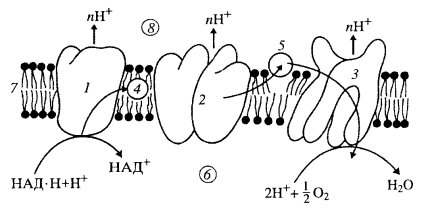
Рис.3 . Поток электронов через три главных ферментативных комплекса при переносе электронов от НАД·Н к О2
1 — НАД·Н-дегидрогеназный комплекс; 2 — с1-комплекс; 3 — цитохромоксидазный комплекс; 4 — убихинон; 5 — цитохром с; 6 — матрикс митохондрии; 7 — внутренняя митохондриальная мембрана; 8 — межмембранное пространство
Комплекс 1 катализирует перенос двух электронов от НАДН к убихинону и он также способен транспортировать через мембрану протоны. Комплекс 1 содержит ФМН и несколько белков с железносерными центрами с типом кластера 2 Fe – 2S и 4 Fe – 4S (рис.4). Согласно современным представлениям, эти центры представляют собой многоядерные комплексы железа, ковалентно связанные с атомами серы и цистеиновыми остатками белков .
Рис.4. Схема расположения железосерных кластеров в белковых молекулах. (Костюк, 321)
Комплекс 11 катализирует окисление сукцината убихиноном и не может переносить протоны. Комплекс содержит ФАД, цитохром b557 и два железосерных центра Fe – S с типом кластера 2 Fe – 2S и один железносерный центр с типом кластера 4 Fe – 4S. Убихинон (коэнзим Q) в митохондриях животных клеток представлен в форме Q10, т.е. его молекула содержит цепь из 10 пятиуглеродных изопреновых остатков, что определяет высокую гидрофобность этой молекулы. Убихинон хорошо растворяется в липидной фазе мембраны и способен к трансмембранному переходу путем диффузии. Присоединяя два электрона и два протона, убихинон восстанавливается и превращается в убихинол QН2.
Комплекс 111 катализирует перенос электронов от убихинола к цитохрому с, способен также транспортировать протоны. В состав этого комплекса входят цитохромы b562 , b566 , цитохром c1, железносерный центр с типом кластеров 2 Fe – 2S.
Комплекс 1У переносит электроны от цитохрома с к кислороду. В состав этого комплекса входят фермент цитохром с-оксидаза с двумя гемами в молекулах цитохромов а и а3 и два атома меди. Окисленная форма (Fe3+) цитохромов а и а3 принимает электроны от восстановленного цитохрома с и переходит в восстановленную форму (Fe2+).
Значительная часть свободной энергии, которая выделяется при переносе электронов по ЭТС, запасается в форме АТФ на участках сопряжения биологического окисления с фосфорилированием. Участки сопряжения локализованы: 1) между НАДН-дегидрогеназой и цитохромом b, 2) между цитохромом b и цитохромом с, 3) между цитохромом с и кислородсвязывающим участком комплекса 1У (см. рис. 3). На этих участках ЭТС количество выделяющейся энергии достаточно для образования фосфоангидридной связи.
По-существу, транспорт электронов по дыхательной цепи митохондрии представляет собой последовательнось окислительно-восстановительных реакций. Перенос электронов в ЭТС осуществляется по градиенту значений окислительно-восстановительных потенциалов (ОВП) пар: донор электронов-акцептор электронов. Как известно, значение ОВП выражается в вольтах и характеризует способность донора отдавать электрон(ы) акцептору в сопряженной окислительно-восстановительной паре. ОВП окислительно-восстановительной пары может иметь и положительное, и отрицательное значение. Чем выше абсолютное значение ОВП системы, тем выше окислительно-восстановительные свойства системы. Величина и знак ОВП для окислительно- восстановительных пар позволяет предсказать направление движения электронов: электроны транспортируются от отрицательно заряженного донора к положительно заряженному (по отношению к донору) акцептору. Определение ОВП окислительно-восстановительных пар проводят путем измерения электродвижучей силы (ЭДС) с использованием стандартного (водородного) электрода. Водородный электрод представляет собой платиновую пластину, который погружен в 1 М раствор Н+ находящийся в равновесии с газообразным Н2 при давлении 1 атм. Потенциал такого стандартного электрода условно принимают равным нулю. Стандартный окислительно восстановительный потенциал Е0 характеризует способность исследуемой редокс-системы окислять или восстановливать водородный электрод, при однаковой концентрации окислителя и восстановителя. Величину ОВП можно найти по следующей формуле
Е0 = Е0 + RTln Ox/ Red/nF
где Е0 – стандартный редокс-потенциал; R – универсальная газовая постоянная; Т- абсолютная температура; Ox/ Red –отношение концентрации окисленной и восстановленной форм вещества; F- число Фарадея; n –число переносимых электронов.
В электронно-транспортной цепи митохондрий восстановителем (донором электронов) является атом водорода ( в составе НАДН и ФАДН2), окислителем служит атом кислорода, который в процессе дыхания восстановливается до Н2О. Как отмечалось выше, между этими молекулами находятся не менее 10 промежуточных молекулярных переносчиков электронов, за счет работы которых энергия окислительно-восстановительных реакций преобразуется в энергию фосфоангидридной связи в молекуле АТФ. Энергия образования фосфоангидридной связи в АТФ при стандартных условиях составляет ΔG0 = - 35 кДж/моль, что соответствует изменению редокс-потенциала Δ Е0 = - ΔG0/ nF = 0,18 В. Величина окислительно–восстановительного потенциала ЭТС митохондрий составляет 1,14 В, что эквивалентно 220 Кдж. В таблице 1 приведены Значения ОВП окислительно-восстановительных пар (основных молекулярных переносчиков), локализованных в дыхательной цепи.
Таблица 1.
Значения окислительно-восстановительных потенциалов молекулярных переносчиков электронов в дыхательной цепи митохондрий
| Донор электронов (восстановленная форма) | Акцептор электронов (окисленная форма) | Е0, В |
| НАДН ФМНН2 ФАДН2 Убихинон (КоQ Red ) Цитохром b (Fe2+) Цитохром c1 (Fe2+) Цитохром c (Fe2+) Цитохром a (Fe2+) Цитохром a3 (Fe2+) Н2О | НАДН+ ФМН ФАД Убихинол (КоQ Ox ) Цитохром b (Fe3+) Цитохром c1 (Fe3+) Цитохром c (Fe3+) Цитохром a (Fe3+) Цитохром a3 (Fe3+) ½ О2 + 2Н+ | - 0,32 - 0,12 - 0,06 0,1 0,07 0,21 0,25 0.29 0,38 0,82 |
Синтез АТФ на мембранах митохондрий. Фосфорилирование АДФ осуществляется встроенным в внутренную мембрану митохондрий ферментом - Н+ -зависимой АТФ-синтазой. По современным представлениям, мембранная АТФ-синтаза является миниатюрным молекулярным мотором. Принцип его работы подобен работе электродвигателя, т.е. вращение ротора происходит вследствие прохождения электрического тока через его обмотку. В отличие от искусственных электромоторов, в АТФ-синтазе вращение ротора осуществляется не потоком электронов, а потоком протонов. До недавнего времени считалось, что самыми миниатюрными «живыми» моторами являются флагеллярные моторы, которые генерируют движение бактерий. Однако, оказалось, что самым маленьким из всех известных в природе вращающихся моторов является протонная АТФ-синтаза. Такие молекулы обнаружены в митохондриях, хлоропластах, на плазматических мембранах бактерий. Исследования последних лет показали, что АТФ-азы в энергопреобразующих мембранах клеток растений, животных и бактерий по струтуре и функциям не имеют существенных различий.
На рис. 4 показана структура ферментного комплекса, полученная на основе данных рентгеноструктурного анализа АТФ-азы из митохондрий сердца быка. Как видно, молекула этой АТФ-азы состоит из двух функциональных частей: встроенного в мембрану протонного канала F0 и каталитической части F1, выступающей в матрикс митохондрии. Этот ансамбль имеет вид слегка приплюснутого шара высотой 8 нм и шириной 10 нм. В центре шара находится субъединица g, которая образована двумя протяженными полипептидными цепями и напоминает слегка деформированный изогнутый стержень длиной около 9 нм. Нижняя часть субъединицы g выступает из шара на 3 нм в сторону мембранного комплекса F0. Субъединица d расположена на внешней стороне F1. Внутри ансамбля (ab)3 находится минорная субъединица e , которая связана с субъединицей g. Обе эти субъединицы (g и e ) подвижны — они входят в состав своеобразного ротора, который вращается внутри неподвижного комплекса (ab)3 .
Мембранный комплекс F0 служит основанием, которое удерживает АТФ-синтазу в мембране.Этоткомплекс включает в себя протонный канал, по которому ионы водорода переносятся через АТРсинтазу Пространственная структура F0 расшифрована не столь детально, как строение водорастворимого комплекса f1. Ориентированный в водную фазу (в матрикс митохондрии) комплекс F1 состоит из девяти субъединиц пяти типов полипептидов (Зa, Зb, g, d, e ). Полипептидные цепи субъединиц a и b уложены в похожие по строению белковые глобулы, которые все вместе образуют гексамер -ансамбль, состоящий из шести субъединиц.
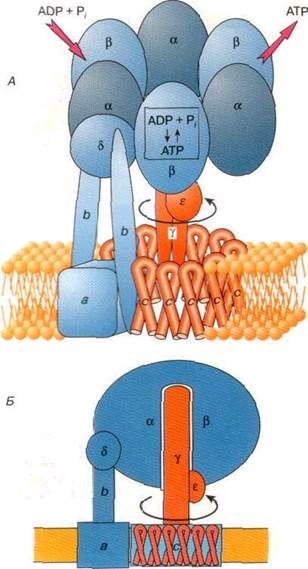
Рис. 4. Схематичное изображение АТФ-синтазы в мембранах митохондрий
А. Схема расположения основных белковых субьединиц, образующих комплексы F0 и F1
Б. АТФ-синтаза - внутриклеточный электродвигатель: Ротор – g, e - субьединицы (обозначены красным цветом); статор - Зa, Зb, d, c, a, b - субьединицы (обозначены синим цветом).
Представления об АТФ-синтазе, как молекулярной машине, работа которой связана с ее вращением, хорошо согласуются со структурными особенностями комплекса. В структуре АТФ-синтазы можно выделить две группы белковых субъединиц: одна из них образует статор мотора, который неподвижен относительно мембраны, а другая соответствует подвижному ротору, вращающемуся внутри статора. Статор включает в себя шарообразный гексамер, состоящий из трех субъединиц a и трех субъединиц b, находящуюся на его поверхности субъединицу d, а также субъединицы a и b мембранного комплекса F0 (рис 4,Б). В этой макромолекулярной конструкции субъединицы b выполняют роль своеобразного кронштейна, связывающего неподвижные субъединицы комплексов F0 и F1. К находящейся в мембране субъединице а примыкает гидрофобное кольцо, образованное субъединицами с мембранного комплекса F0.
Ротор состоит из субъединиц g и e комплекса F1. Субъединица g, расположенная внутри комплекса (ab)3 , заметно выступает из него и соединяется с погруженным в мембрану кольцом из субъединиц с. Имеются все основания считать, что субъединица g, входящая в состав ротора, действительно вращается при работе фермента. Для того, чтобы провернуть ротор внутри статора, и тем самым заставить АТФ- синтазу сделать молекулу АТР, необходим внешний источник энергии. Как уже было сказано выше, когда АТФ-синтаза работает в режиме синтеза АТФ, движущей силой для ее работы является энергия протонов, переносимых через сопрягающую мембрану за счет протонного потенциала. При работе АТФ-синтазы в режиме гидролиза АТФ источником энергии для вращения ротора служит энергия, запасенная в молекуле АТФ.
Наглядно показано, что гидролиз АТР комплексом F1, действительно сопровождается вращением субъединицы g относительно гексамера (ab)3. Об этом свидетельствуют работы американцев Капальди, Кросса и их сотрудников. Им удалось пришить субъединицу g к субъединице b и тем самым блокировать возможное вращение субъединицы g внутри комплекса F1. В этом случае, ферментативная активность комплекса F1, (его способность гидролизовать АТФ) при этом была полностью подавлена.
Самым впечатляющим доказательством того, что субъединица γ, действительно крутится в ходе работы фермента, стала замечательная работа, группой японских исследователей Киношите, Йошиде и их соавторами. Они увидели вращение субъединицы γ, с помощью флуоресцентного микроскопа. Как можно разглядеть вращение ротора, диаметр которого составляет всего лишь 1 нм ? Чтобы наблюдать за вращением этой субъединицы, к ее основанию, выступающему из комплекса F1, японские ученые прикрепили специальный макромолекулярный маркер - фрагмент нити актина длиной около одного микрона, который, в свою очередь, был помечен флуоресцентным красителем. Остальную часть отделенной от мембраны молекулы f1 обездвижили, пришив к субъединицам b специальные хвостики, с помощью которых F1 прикрепили к неподвижной подложке. Наблюдая с помощью микроскопа за изменением положения флуоресцирующей нити актина, жестко связанной субъединицей γ, удалось непосредственно увидеть ее вращение. Оказалось, что в ходе работы фермента, гидролизующего АТФ, актиновый хвост вращается против часовой стрелки. Эти эксперименты продемонстрировали, в буквальном смысле этого слова, вращение самого маленького из всех известных к настоящему времени природных моторов. Вместе с этим в науке окончательно утвердилось новое понятие — вращательный катализ (англ. — rotary catalysis).
Замечательным качеством вращающегося мотора АТФ-синтазы является его исключительно высокий коэффициент полезного действия. Показано, что для поворота актинового хвоста на 120° затрачивается около 35 кДж энергии, т.е. приблизительно равная энергии образования АТФ из АДФ. Это означает, что КПД работы мотора близок к 100 %.
В табл. 2 приведены сравнительные характеристики различных молекулярных моторов, встречающихся в живой клетке. Видно, что АТФ-синтаза является своего рода рекордсменом среди молекулярных моторов своей "весовой категории". По эффективности работы и развиваемой ею силе она существенно превосходит все известные в природе молекулярные моторы. Так, например, максимальная сила, создаваемая при работе одного миозинового мостика актомиозинового комплекса мышечных волокон, составляет Fмакс = 3-5 пН. Вращательный момент, создаваемый молекулой f1 за счет гидролиза АТФ, достигает величины М = 40 пН • нм. Если учесть, что радиус вращающейся субъединицы составляет r = 1 нм, то сила Fмакс развиваемая молекулой f1, будет равна 40 пН. Оказывается, что молекула F1, приблизительно в 10 раз сильнее актомиозинового комплекса - молекулярной машины, специализирующейся в клетках и различных органах на "профессиональном" выполнении механической работы. Таким образом, за сотни миллионов лет до того, как появился человек, который изобрел колесо, преимущества вращательного характера движения были успешно реализованы природой на молекулярном уровне.
Таблица 2.
Дата добавления: 2018-03-02; просмотров: 4561;