П - показатель степени (для большинства рецепторов около 1).
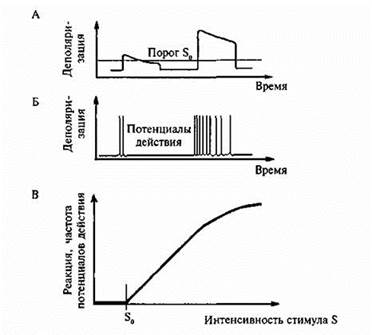
Рис. 3. Степенная функция рецептора.
А -генераторный потенциал рецептора;
Б - потенциалы действия в афферентном волокне;
В - график зависимости частоты импульсации в афферентном волокне от силы стимула;
S - величина стимула;
S0- пороговый стимул.
Большое число рецепторов спонтанно разряжаются в отсутствие каких - либо раздражителей. Например, рецепторы вестибулярного аппарата постоянно активны, что дает им возможность сигнализировать не только о величине, но и о направлении действующего стимула (например, ускорения). Для этих случаев порог определяется не как возникновение, а как изменение реакции рецептора - дифференциальный порог.
5. При длительном действии адекватного стимула порог срабатывания данного рецептора повышается. Это явление называется адаптацией. В адаптации рецептора принимает участие большое число процессов, и ее нельзя объяснить каким - либо одним механизмом.
Например, в рецепторах кожи самым важным местом адаптации является ткань, окружающая сенсорное нервное окончание. К этой механической адаптации прибавляется адаптационный процесс генераторного потенциала.
В состав сенсорной системы, кроме описанного 1 -го уровня обработки информации, входит также ряд других структур головного мозга с соответствующими проводящими путями. Одними из важных станций переключения афферентной импульсации к коре служат специфические ядра таламуса. Высшим уровнем обработки сенсорных сигналов является кора больших полушарий, которая достигает наибольшего развития у млекопитающих и особенно у приматов.
Толщина коры у млекопитающих разных видов колеблется от 1 до 6 мм. Нейроны по толщине коры распределены неравномерно и обычно образуют в новой коре 6 слоев, в старой коре - 3 слоя (рис.5). Однако в отдельных областях новой коры количество слоев может увеличиваться или уменьшаться. Слои V и VI содержат преимущественно крупные пирамидные нейроны, аксоны которых образуют эфферентные пути из коры. Основные афферентные пути в кору заканчиваются на нейронах верхних слоев (III и IV). Эти слои наиболее сильно развиты в центральных отделах зрительного, слухового и кожного анализаторов. По современным представлениям, такое деление слоев коры на афферентные и эфферентные нужно считать в значительной степени условным. В последние годы при изучении внутрикорковых связей установлено, что эфферентные аксоны нейронов нижних слоев образуют многочисленные возвратные коллатерали, которые восходят до самых верхних слоев коры.
-

В соответствии с цитоархитектоническими и нейрофизиологическими данными выделяют проекционные и ассоциативные зоны коры. В проекционных зонах различают первичные, вторичные и третичные зоны. По И. П. Павлову, под ядром анализатора понимают зону коры, в которой после соответствующих переключений в подкорковых ядрах имеет место детальное представительство периферического отдела анализатора. Принцип деления на первичные, вторичные и третичные поля иллюстрируется рис.6, на котором схематично показано, что возбуждение от соответствующих рецепторов направляется в первичные зоны по быстропроводящим путям, тогда как активация вторичных и ассоциативных зон коры происходит по полисинаптическим путям. Кроме этого, корковые поля связаны между собой многочисленными ассоциативными волокнами. В корковых проекциях сенсорных систем соблюдается принцип топии, например, в зрительной системе рети нотопия - проекция сетчатки - в поле 17 (первичная проекция), поле 18 (вторичная) и поле 19 (третичная). Этот принцип состоит в том, что, например, сетчатка образует проекцию на соответствующую область коры в некотором масштабе. При этом объем корковой проекции пропорционален плотности рецепторов в данном месте сетчатки. Благодаря этому fovea (зрительная ямка) сетчатки в корковой проекции представлена большей площадью, чем периферия сетчатки. Поражение первичных зон сенсорных систем всегда сопровождается грубыми нарушениями соответствующей сенсорной или моторной функции. В отличие от этого поражение вторичных или третичных зон вызывает расстройство аналитико - синтетической деятельности мозга, например гнозиса и праксиса, речи, целенаправленного поведения и пр.
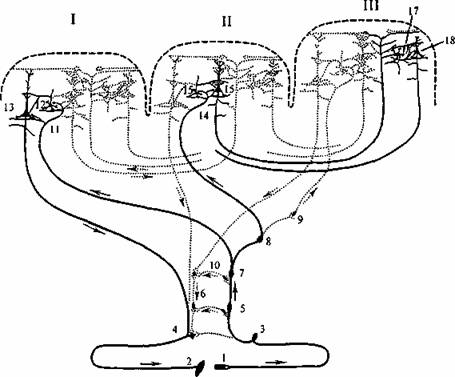
Рис. 6.6. Система связей первичных, вторичных и третичных полей коры (по Полякову, 1965).
I первичные (центральные) поля; II - вторичные (периферические) поля; III - третичные (ассоциативные) поля (зоны перекрытия анализаторов); жирными линиями выделены: система проекционных (корково -подкорковых) связей коры; система проекционно - ассоциативных связей коры; система ассоциативных связей коры; 1 - рецептор; 2 - эффектор; 3 - нейрон чувствительного узла; 4 - двигательный нейрон; 5-6 - переключательные нейроны спинного мозга и ствола; 7-10 - переключательные нейроны подкорковых образований; 11, 14 - афферентное волокно из подкорки; 13 - пирамида Услоя; 16 - пирамида подслоя III3; 18 - пирамиды подслоев III2, НИ; 12, 15, 17 - звездчатые клетки коры.
Симультанное и сукцессивное восприятие.
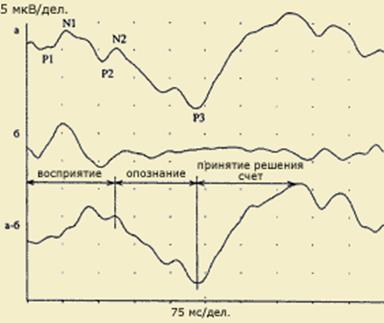
Все виды восприятия несут информацию о времени, т.е. о том моменте, когда появился стимул и как долго он действовал. Другими словами, восприятие - это процесс, началом которого служит момент действия стимула на рецептор. Завершением восприятия является образ стимула - объекта и его опознание (идентификация). Длительность одного акта восприятия может быть очень короткой, почти мгновенной, особенно, когда объект восприятия хорошо знаком. В таком случае говорят об одномоментном (симультанном) восприятии. Если человек сталкивается с неизвестным стимулом, длительность восприятия может существенно увеличиваться. Требуется время, чтобы провести детальный сенсорный анализ, выдвинуть и проверить несколько гипотез по поводу действующего стимула, и лишь после этого принять решение о том, что же представляет собой воспринимаемый стимул. В этом случае говорят о последовательной обработке информации и сукцессивном восприятии.
Изучение физиологических основ восприятия проводится с помощью макропотенциалов мозга: фоновой электроэнцефалограммы, а также вызванных и событийно-связанных потенциалов, причем использование ЭЭГ имеет два разных аспекта. Первый связан с выявлением параметров ЭЭГ, имеющих отношение к кодированию стимула. Второй, более традиционный, представляет собой изучение параметров ЭЭГ отдельных зон коры или показателей их совместного взаимодействия при восприятии стимулов.
Среди ритмов ЭЭГ наибольшее внимание исследователей в этом плане привлекает альфа-ритм (8-12 кол/с), который регистрируется преимущественно в задних отделах коры в состоянии спокойного бодрствования. Известно, что при предъявлении стимулов имеет место подавление или "блокада" альфа-ритма: причем она тянется тем больше, чем сложнее изображение.
Предполагается, что такая особенность ЭЭГ, как пакет волн создается синхронизированной когерентной активностью группы нейронов, расположенных в разных участках мозга и образующих ансамбль. Предположительно все волны одного пакета хранят информацию об одном образе или его части и при восприятии опознается только тот образ, который закодирован ритмической активностью наибольшего числа нейронов в каждый данный момент времени.
Наряду с этим существуют исследования пространственно-временных отношений потенциалов мозга при восприятии сенсорной информации. В отличие от предыдущего этот подход учитывает два фактора: время восприятия и его мозговую организацию.
Реорганизация биоэлектрической активности в процессе перцептивного акта характерна для всех видов чувствительности. Однако в зависимости от вида анализатора она имеет разную пространственную картину. Как правило, на ранних этапах процесса наибольшую активность демонстрирует проекционная зона (зрительная, слуховая, соматосенсорная), на завершающих этапах в процесс включаются передние отделы коры.
 Наиболее адекватным инструментом изучения физиологической природы перцептивного процесса является метод регистрации вызванных (ВП), или событийно-связанных потенциалов. Последнее обусловлено тем, что, во-первых, ВП - это реакция мозга на внешнее раздражение, по длительности сопоставимая со временем перцептивного акта, во-вторых, ВП, взятые отдельно в каждой зоне коры, имеют свои характеристики, обусловленные функциональными особенностями этой зоны. Таким образом, при предъявлении стимула любой модальности (зрительного, звукового, тактильного и др.) в разных зонах коры возникают специализированные реакции, сопоставимые по длительности с реальным психологическим процессом восприятия.
Наиболее адекватным инструментом изучения физиологической природы перцептивного процесса является метод регистрации вызванных (ВП), или событийно-связанных потенциалов. Последнее обусловлено тем, что, во-первых, ВП - это реакция мозга на внешнее раздражение, по длительности сопоставимая со временем перцептивного акта, во-вторых, ВП, взятые отдельно в каждой зоне коры, имеют свои характеристики, обусловленные функциональными особенностями этой зоны. Таким образом, при предъявлении стимула любой модальности (зрительного, звукового, тактильного и др.) в разных зонах коры возникают специализированные реакции, сопоставимые по длительности с реальным психологическим процессом восприятия.
В конфигурации ВП можно выделять компоненты двух
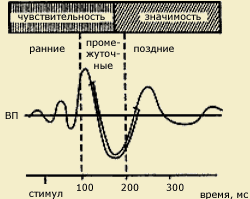
| Вызванные потенциалы (ВП) мозга, возникающие в ответ на световой или кожный раздражитель (по А.М.Иваницкому, 1999). Видно, что ранние волны соотносятся с чувствительностью, поздние - с мотивацией, значимостью стимула, а промежуточные - одновременно с двумя этими психофизическими показателями. Стрелкой на временной шкале обозначает момент предъявления стимула |
типов: ранние специфические (экзогенные) и поздние неспецифические (эндогенные) компоненты. Обработка физических параметров стимула (сенсорный анализ) связывается с деятельностью специфических сенсорных систем и находит свое отражение преимущественно в параметрах экзогенных компонентов ВП. Эндогенные компоненты ВП, возникающие позднее и длящиеся до окончания ответа, предположительно отражают этапы более сложной обработки стимула: формирование образа, сличение его с эталонами памяти, принятие перцептивного решения.
Дата добавления: 2017-12-05; просмотров: 539;