Интегративные феномены в ЦНС.
Посттетаническая потенциация. Раздражая афферентный нерв стимулами редкой частоты, можно получить некоторый рефлекс определенной интенсивности. Если затем этот нерв в течение некоторого времени подвергать высокочастотному раздражению (300 – 400 стимулов в с), то повторное редкое ритмическое раздражение приведет к резкому усилению реакции.
Окклюзия (закупорка). Если 2 нервных центра рефлекторных реакций имеют частично перекрываемые рецептивные поля, то при совместном раздражении обоих рецептивных полей реакция будет меньше, чем арифметическая сумма реакций при изолированном раздражении каждого из рецептивных полей. Прим., 1-й нейрон активирует 10 мышечных волокон, развивается мышечное напряжение 100 мгс. 2-й нейрон возбуждает также 10 других волокон (100 мгс). Но если оба нейрона возбуждать одновременно, то суммарная активность мышцы 180 мгс. Часть волокон были общими (т.е. нейрон 1 и 2 передавали информацию на одни и те же волокна).
Облегчение. В ряде случаев при совместном раздражении рецептивных полей двух рефлексов вместо ослабления (окклюзии) наблюдается облегчение (т.е. суммарная реакция выше суммы реакции при изолированном раздражении этих рецептивных полей). Часть общих для обоих рефлексов нейронов при изолированном раздражении оказывают подпороговый эффекты. При совместном раздражении они суммируются и достигают пороговой силы.
Торможение.
Торможение в ЦНС – активный процесс, проявляющийся в подавлении или в ослаблении процесса возбуждения. Т.е. в норме торможение является производным от возбуждения, ограничивая и препятствуя его чрезмерному распространению. Процесс торможения вместе с процессом возбуждения формируют сложную мозаику активированных и заторможенных зон в центральных нервных структурах.
История развития учения о тормозных процессах начинается с открытия И.М. Сеченовым центрального торможения. Химическое раздражение кристаллами соли зрительных бугров вызывало удлинение времени сгибательного рефлекса по методике Тюрка (сгибание конечности в тазобедренном и коленном суставах в ответ на погружение лапы в кислоту).
Гольц получил «периферическое торможение», применяя дополнительные раздражители периферии (конечности).
Возникло предположение о существовании специфических тормозных нейронов, оказывающих тормозные влияния на другие нейроны. Впоследствии было показано (Экклс, Реншоу) существование специальных вставочных нейронов, имеющих синаптические контакты с другими нейронами и оказывающих на них тормозное влияние. Некоторые из этих клеток имеют «собственное» имя, например клетки Реншоу. Результатом деятельности тормозных нейронов может быть не только торможение нейронов. В коре и гиппокампе есть тормозные нейроны, вызывающие торможение других тормозных нейронов и, вследствие этого, растормаживающий эффект.
Если рассмотреть «архитектуру» использования тормозных нейронов в организации нейронных цепей, то можно выделить ряд вариантов:
1) Реципрокноеторможение. Пример: сигнал от мышечного веретена поступает с афферентного нейрона в спинной мозг, где переключается на альфа-мотонейрон сгибателя и одновременно на тормозной нейрон, который тормозит активность альфа-мотонейрона разгибателя (Ч. Шеррингтон);
2) Возвратное(антидромное)торможение. Угнетение активности нейрона вызывается возвратной коллатералью аксона, заканчивающейся на тормозном нейроне, аксон которого оказывает тормозное действие. Пример: альфа-мотонейрон посылает аксон к соответствующим мышечным волокнам. По пути от аксона отходит коллатераль, которая возвращается в ЦНС – она заканчивается на тормозном нейроне (клетка Реншоу) и активирует ее. Тормозной нейрон вызывает торможение альфа-мотонейрона, который запустил всю эту цепочку. Таким образом, альфа-мотонейрон, активируясь, через систему тормозного нейрона тормозит сам себя;
3) Латеральноеторможение (вариант возвратного торможения). Пример: Фоторецептор активирует биполярную клетку и, одновременно, рядом расположенный тормозной нейрон, блокирующий проведение возбуждения от соседнего фоторецептора к ганглиозной клетке («вытормаживание информации» - 2 точки на сетчатке рассматриваются как раздельные точки, если между ними есть невозбужденные участки).
Механизмы торможения.
Различают несколько видов торможения: постсинаптическое, пресинаптическое, пессимальное (рис. 16).
Постсинаптическоеторможение – развивается на ПСМ аксосоматических и аксодендритических синапсов под влиянием тормозных нейронов, у которых из концевых разветвлений аксонных отростков в синаптическую щель высвобождается тормозной медиатор (например, ГАМК или глицин). Медиатор вызывает на ПСМ эффект гиперполяризации в виде ТПСП, а пространственно-временная суммация ТПСП приводит к урежению или прекращению генерации ПД в результате снижения возбудимости нейрона. Это основной вид торможения.
Пресинаптическоеторможение – развертывается в аксоаксональных синапсах, блокируя распространение возбуждения по аксону. Процесс торможения здесь протекает по типу катодической депрессии: в области контакта выделяется медиатор (ГАМК), который вызывает стойкую деполяризацию, что нарушает проведение волны возбуждения через этот участок. Является наиболее избирательным видом торможения, т.к. выключает отдельные входы к нервной клетке.
Блокатором ГАМК-ергических рецепторов мембраны является бикукулин, а глициновых рецепторов – стрихнин и столбнячный токсин.
Эти два вида торможения, возникающие в результате деятельности тормозных нейронов, относят к первичному торможению (Дж. Экклс).
Пессимальное(вторичное) торможение – вид торможения центральных нейронов, наступает при высокой частоте раздражения. В первый момент возникает высокая частота ответного возбуждения, однако через некоторое время стимулируемый в таком режиме нейрон переходит в состояние торможения.
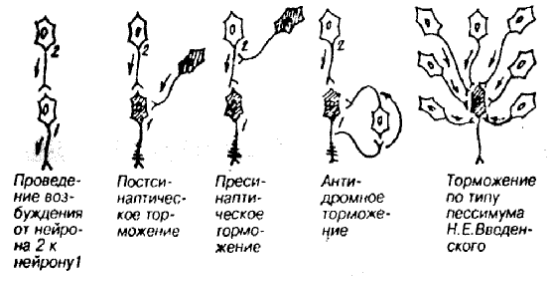
Рис. 16. Виды внутрицентрального торможения.
2.2.4. Принципы координации в деятельности ЦНС.
Конвергенция (концентрация). Ч. Шеррингтон называл этот принцип - «принципом общего конечного пути». Многие нейроны оказывают свое воздействие на один и тот же нейрон, т.е. имеет место схождение потоков импульсов к одному и тому же нейрону. Прим., сокращение мышцы (за счет возбуждения альфа-мотонейрона) можно вызвать за счет растяжения мышцы (рефлекс мышечных веретен) или путем раздражения кожных рецепторов (сгибательный рефлекс).
Дивергенция. Способность нервной клетки устанавливать многочисленные синаптические связи с различными нервными клетками. Обеспечивает иррадиацию возбуждения в центральных нервных образованиях. Тормозные процессы ограничивают дивергенцию и делают процессы управления более точными. Когда торможение снимается (например, при столбняке), происходит полная дискоординация в деятельности ЦНС.
Принцип реципрокной иннервации (см. – реципрокное торможение).
Принцип обратной связи и копий эфферентаций. Невозможно точно координировать, управлять, если отсутствуют данные о результатах управления. Осуществляется за счет потока импульсов с рецепторов. Копия эфферентации: Для управления важно иметь информацию о командах, посылаемых на периферию. Например, в системах управляющих скелетными мышцами, каждый отдел, посылая сигнал управления к работающей мышце, одновременно сообщает об этом вышележащему отделу (вариант обратной связи).
Принцип доминантыоткрыт А.А.Ухтомским. Изучая ответы скелетной мышцы кошки на электрические раздражения КБП, он обнаружил, что при акте дефекации ответы мышцы прекращаются. Он пришел к выводу, что среди рефлекторных актов, которые могут быть выполнены в данный момент времени, имеются рефлексы, реализация которых является в данный момент времени важнейшей для организма. Центры, участвующие в реализации доминантных рефлексов он назвал «доминантным очагом возбуждения». Свойства доминантного очага: он стойкий (его трудно затормозить); интенсивность его возбуждения усиливается слабыми раздражителями; этот очаг тормозит другие потенциальные доминантные очаги. Доминантность того или иного очага определяется состоянием организма (у голодного животного доминируют пищевые рефлексы).
Принцип системности. Развитием представления о доминанте являются работы П.К. Анохина о функциональной системе.
Рефлекторные реакции протекают не изолированно, а всегда объединяются в систему. Любая функциональная система всегда формируется и функционирует для достижения организмом конкретных приспособительных результатов (главный системообразующий фактор – конечный результат действия). Любая рефлекторная реакция многокомпонентная и протекает в 4 стадии.
1 стадия – афферентный синтез. На организм действует огромное количество раздражителей – это обстановочная афферентация. Есть аппарат памяти – прошлый опыт, аппарат мотиваций – побуждение к деятельности. Происходит их суммация и выделяется главный раздражитель – пусковая афферентация.
2 стадия – принятие решения. На организм действуют те же раздражители: возбужден аппарат памяти, мотиваций, выделена пусковая афферентация. Происходит принятие решения, это переломный момент в любой рефлекторной реакции.
3 стадия – эфферентный синтез. Возбужден аппарат памяти, мотиваций, выделена пусковая афферентация, принято решение. Формируется 2 функциональных аппарата: 1) программа действия - что, как и в какой последовательности нужно делать для достижения конечного приспособительного результата; 2) акцептор результата действия (АРД) - является аппаратом предвидения, предугадывания. Формируется на основе решения и программы действия. Стадия заканчивается началом действия.
4 стадия – обратной афферентации. АРД является аппаратом сравнения задуманного с полученным. Параметры результата действия поступают в АРД, который необходим для контроля и исправления ошибок в наших действиях. Совпадение задуманного с полученным сопровождается положительными эмоциями, при несовпадении – отрицательными эмоциями. В последнем случае вносятся поправки и система запускается повторно.
Принцип пластичности. При повреждении отдельных центров мозга их функция может перейти к другим структурам мозга (если не связано с наступлением смерти, как при нарушении дыхательного центра). Процесс возмещения утраченных функций осуществляется при обязательном участии КБП.
Принцип иерархичности. Принцип субординации или соподчинения. Подчинение низших отделов нервной системы высшим (филогенетически ранних более поздним). Цефализация нервной системы и ее управляющей функции (проявляется в перемещении, сосредоточении функции регуляции и координации деятельности организма в головных отделах ЦНС). Высшее проявление – кортикализация функций. В КБП имеются нейронные комплексы (ансамбли), отвечающие за все функции организма. Высшие нервные центры выступают уже регуляторами регуляторов. При всей сложности взаимоотношений между старыми, древними и эволюционно новыми образованиями мозга общая схема следующая: восходящие влияния (от нижележащих старых структур к вышележащим новым) преимущественно возбуждающего, стимулирующего характера, а нисходящие - преимущественно угнетающего, тормозного характера. Т.е. в процессе эволюции повышается роль и значенияетормозных процессов в осуществлении сложных интегративных рефлекторных реакций.
Принцип иерархичности проявляется и в общей закономерности расположения нейронов в ЦНС:
- спинной мозг – «ядра» на всем протяжении;
- продолговатый – крупные ядра;
- гипоталамус – много мелких ядер;
- КБП – слои нервных клеток.
Чем сложнее функция, тем упорядоченнее расположение нервных клеток. Благодаря принципу иерархичности:
1) Расширяются возможности целостного организма, возможна более тонкая, дифференцированная регуляция функций;
2) Повышается коррекция результатов деятельности многих органов, в том числе и анализаторов.
Принцип целостности. Органически сочетается с принципами иерархичности и системности. Подразумевает функционирование всех звеньев или этажей ЦНС.
СПИННОЙ МОЗГ
Характерной чертой организации спинного мозга (СМ) является периодичность его структуры в форме сегментов, имеющих входы в виде задних корешков, клеточную массу нейронов (серое вещество) и выходы в виде передних корешков.
СМ имеет 31- 33 сегмента: 8 шейных (С1-С8), 12 грудных (Т1- Т12), 5 поясничных (I1-I5), 5 крестцовых (S1-S5), 1-3 копчиковых (Со1-Со3).
Деление на сегменты является больше функциональным, чем морфологическим и определяется зоной распределения в нем волокон заднего корешка и зоной клеток, которые образуют выход передних корешков. Каждый сегмент через свои корешки иннервирует 3 метамера тела и получает информацию также от 3 метамеров. В итоге перекрытия каждый метамер тела иннервируется 3-мя сегментами и передает сигналы в 3 сегмента СМ.
В опытах с перерезкой и раздражением корешков спинного мозга показано, что задние корешки являются афферентными, чувствительными, центростремительными, а передние – эфферентными, двигательными, центробежными (закон Белла – Мажанди).
Афферентные входы в спинной мозг организованы аксонами спинальных ганглиев, лежащих вне спинного мозга, и аксонами экстра- и интрамуральных ганглиев симпатического и парасимпатического отделов АНС.
1-я группа афферентных входов образована волокнами, идущими от мышечных рецепторов, рецепторов сухожилий, надкостницы, оболочек суставов (начало проприоцептивной чувствительности); 2-я группа входов начинается от кожных рецепторов (кожная рецептирующая система); 3-я – висцерорецептивная система.
Серое вещество в шейном, поясничном и крестцовом отделах СМ образовано его нейронами в виде симметрично расположенных 2-х передних и 2-х задних рогов. В грудном отделе есть еще и боковые рога.
Задние рога выполняют главным образом сенсорные функции и содержат нейроны, передающие сигналы в вышележащие центры, в симметричные структуры противоположной стороны либо к передним рогам СМ.
В передних рогах находятся эфферентные нейроны, дающие свои аксоны к мышцам. Все нисходящие пути ЦНС, вызывающие двигательные реакции, заканчиваются на нейронах передних рогов («общий конечный путь» по Шеррингтону).
С 1-го грудного сегмента СМ и до первых поясничных сегментов, в боковых рогах серого вещества располагаются нейроны симпатического, а в крестцовых – парасимпатического отдела АНС.
СМ человека содержит 13 млн нейронов, из них 3% - мотонейроны, 97% - вставочные. Функционально делятся на 4 группы:
1) - мотонейроны – клетки передних рогов, аксоны которых образуют передние корешки;
2) - интернейроны – располагаются в задних рогах, получают информацию от спинальных ганглиев, реагируют на болевые, температурные, тактильные, вибрационные, проприоцептивные раздражения;
3) - симпатические и парасимпатические нейроны - расположены преимущественно в боковых рогах. Их аксоны выходят в составе передних корешков;
4) - ассоциативные клетки – нейроны собственного аппарата СМ, устанавливающие связи внутри и между сегментами.
В средней зоне серого вещества (между задним и передним рогами) имеется промежуточное ядро (ядро Кахаля) с клетками, аксоны которых идут вверх или вниз на 1-2 сегмента и дают коллатерали на нейроны ипси- и контрлатеральной стороны, образуя сеть. Подобная сеть имеется и на верхушке заднего рога (студенистое вещество или желатинозная субстанция Роланда) и выполняет функции ретикулярной формации СМ.
Белое вещество СМ состоит из миелиновых волокон, которые собраны в пучки. Эти волокна могут быть короткими (межсегментарные) и длинными.
Короткие волокна (ассоциативные, проприоспинальные) связывают нейроны разных сегментов или симметричные нейроны противоположных сторон СМ.
Длинные волокна (проекционные) делятся на восходящие, идущие к головному мозгу, и нисходящие, идущие от головного мозга. Эти волокна образуют проводящие пути СМ.
Нисходящие пути:
Передний (прямой – спускается до своего сегмента и там переходит на мотонейроны противоположной стороны) и латеральный (перекрещенный – перекрест на уровне продолговатого мозга) корково-спинномозговой (от пирамидных нейронов пирамидной и экстрапирамидной коры, обеспечивает регуляцию произвольных движений);
Красноядерно-спинномозговой (руброспинальный) состоит из аксонов нейронов красного ядра. Переходят на симметричную сторону сразу после выхода из ядра (средний мозг) и делятся на 3 пучка (в СМ, мозжечок, ретикулярную формацию ствола). Участвует в управлении мышечным тонусом;
Преддверно-спинномозговой (вестибулоспмнальный) путь начинается от нейронов латерального преддверного ядра (ядро Дейтерса), лежащего в продолговатом мозге. Регулирует активность мотонейронов СМ, тонус мускулатуры, согласованность движений, равновесие;
Ретикулярно-спинномозговой (ретикулярный) путь идет от ретикулярной формации ствола мозга к мотонейронам СМ, регулирует тонус мускулатуры.
Восходящие пути:
Проприоцептивный (ганглиобульбарный) путь (тонкий и клиновидный пучки) начинается от рецепторов мышц сухожилий, надкостницы, оболочек суставов. Тонкий пучок от ганглиев, собирающих информацию от каудальных отделов тела, таза, нижних конечностей. Клиновидный от ганглиев, собирающих информацию от мышц грудной клетки, верхних конечностей. В тонком и клиновидном ядрах продолговатого мозга переключение на новый нейрон, далее путь в латеральные ядра таламуса противоположного полушария, второе переключение, далее путь к нейронам 4 слоя соматосенсорной области коры. Коллатерали в каждом сегменте СМ для коррекции позы всего туловища. Скорость 60-100 м/с.
Спинно-таламический путь (латеральный и пердний) – начинается от болевых, температурных, тактильных и барорецепторов кожи. Спинальный ганглий, через задний корешок к заднему рогу (первое переключение), аксоны идут на противоположную сторону и поднимаются по боковому канатику к таламусу (2-е переключение), далее – в сенсомоторную область КБП. Часть волокон кожных рецепторов идет к таламусу по переднему канатику. Скорость до 30 м/с.
Спинно-мозжечковые пути лежат в боковых канатиках СМ и представлены неперекрещивающимися передним, спинно-мозжечковым путем (пучок Говерса) и дважды перкрещивающимся задним спинно-мозжечковым путем (пучок Флексига). Т.е. левая доля мозжечка получает информацию с левой стороны тела, а правая с правой. Сухожильные рецепторы Гольджи, проприорецепторы, рецпторы давления, прикосновения. Скорость 110-120 м/с.
При половинном латеральном повреждении СМ развивается синдром Броун-Секара:
На стороне поражения СМ (ниже места поражения) развивается паралич двигательной системы. На противоположной поражению стороне движения сохраняются. Причина – повреждение пирамидных путей. Мотонейроны освобождаются от тормозного влияния пирамидных клеток коры и возникает гипертонус мышц, затем спастический паралич;
На стороне поражения (ниже места поражения) нарушается проприоцептивная чувствительность. Восходящие пути глубокой чувствительности идут по своей стороне СМ до продолговатого мозга (ганглиобульбарный путь);
На противоположной стороне туловища (относительно повреждения СМ) нарушается болевая чувствительность (в спинно-таламических путях переключение в заднем роге на нейрон, аксон которого переходит на противоположную сторону). Если повреждена левая половина СМ, то исчезает болевая чувствительность правой половины туловища ниже повреждения и, наоборот.
Спинальный шок - возникает в случае полного пересечения СМ. Деятельность нейронов СМ на определенное время приостанавливается и все центры ниже перерезки перестают организовывать присущие им рефлексы. Чем сложнее организация ЦНС животного, тем длительней период, в течение которого происходит восстановление функций СМ. У лягушки десятки секунд, у собак отдельные рефлексы восстанавливаются через несколько часов (мышечного сокращения), другие - через несколько дней (рефлексы регуляции артериального давления), или недель (рефлексы мочеиспускания). У человека первые спинальные рефлексы восстанавливаются через несколько недель, а то и месяцев.
Т.е. чем сложнее организация ЦНС у животного, тем сильнее контроль вышележащих отделов мозга над нижележащими. То, что причиной шока является нарушение супраспинальных влияний доказывается повторной перерезкой СМ ниже места первой перерезки. В этом случае спинальный шок не возникает, рефлекторная деятельность СМ сохраняется.
По истечении длительного периода времени после шока спинальные рефлексы резко усиливаются, что объясняется устранением тормозного влияния ретикулярной формации ствола мозга на рефлексы СМ.
При нарушении супраспинальных влияний у человека может появиться группа спинальных рефлексов, которые имеются в норме лишь первые дни и месяцы постнатального развития. Растормаживание этих примитивных рефлексов является клиническим признаком нарушений работы мозга.
Разгибательные и сгибательные патологические рефлексы:
Разгибательные:
Разгибательный рефлекс Бабинского – при раздражении подошвы стопы штриховым движением от пятки до носка происходит разгибание большого пальца в сторону тыла (в норме у детей до 2-х лет).
Рефлекс Оппенгейма – при надавливании на поверхность голени в области в области большеберцовой кости – от колена до голеностопного сустава – происходит тыльное разгибание большого пальца.
Рефлекс Гордона – тыльное разгибание большого пальца происходит при сдавливании икроножной мышцы.
Рефлекс Шефера – то же самое при сдавливании ахиллового сухожилия.
Сгибательные:
Рефлекс Россолимо – сгибание пальцев стопы и в том числе большого при быстром касательном ударе по подушечкам пальцев стопы.
Рефлекс Бехтерева-Менделя – подошвенное сгибание большого пальца при ударе молоточком по тылу стопы.
Рефлекс Жуковского – при ударе по подошвенной поверхности под пальцами стопы.
Рефлекс Бехтерева – при ударе по пятке.
Для верхней конечности:
Сгибательный рефлекс Россолимо – сгибание пальцев кисти в ответ на быстрое касание пальцев;
Рефлекс Жуковского – сгибание пальцев кисти при ударе молоточком по ладонной поверхности кисти;
Рефлекс Бехтерева – при ударе по тылу кисти.
Хватательный рефлекс (Янушкевича) – при касании пальцев руки или стопы проявляется в захватывании предмета (у детей в возрасте до 2-4 месяцев). Иногда при этом удается приподнять ребенка над опорой (хватательный рефлекс Робинсона).
Патологические рефлексы – это по сути истинные рефлексы, которые в процессе онтогенеза заторможены как рудиментарные.
Рефлекторные функции СМ.
Миотатические рефлексы спинного мозга (рефлексы на растяжение мышцы). Рефлекторное увеличение напряжения мышцы в ответ на ее растяжение. Отличаются локальностью и отсутствием существенного последействия. Наиболее выражены в мышцах-разгибателях, которые должны находиться в состоянии тонического напряжения для противодействия силе земного притяжения. Эти рефлексы всегда сопряжены с торможением нервных центров мыщц-антагонистов.
Пример: легкий удар по сухожилию надколенной чашечки вызывает сокращение мышц бедра и разгибание голени. Дуга: мышечные рецепторы четырехглавой мышцы бедра, спинальный ганглий, задние корешки, задние рога 3-го поясничного сегмента, мотонейроны передних рогов того же сегмента, экстрафузальные волокна четырехглавой мышцы бедра.
Рефлексы с рецепторов кожи.Их характер зависит от силы раздражения, вида раздражаемого рецептора, но чаще всего конечная реакция в виде усиления мышц-сгибателей.
Сгибательные рефлексы. Не могут воспроизводиться с большой частотой. Это рефлексы рывкового типа. Усиливаются при перерезке спинного мозга, значит находятся под постоянным тормозным влиянием вышерасположенных нервных центров.
Ритмические рефлексы. Осуществляется сгибание – разгибание. Например, при ходьбе.
Висцеромоторные рефлексывозникают при стимуляции афферентных нервов внутренних органов и характеризуются двигательными реакциями мышц грудной клетки и брюшной стенки, мышц разгибателей спины.
Автономные(вегетативные) рефлексы. Осуществляются за счет нервных центров торако-люмбального и сакрального отделов спинного мозга.
Саморегуляция тонуса скелетных мышц.
Осуществляется при наличии 4 основных образований:
- сухожильные проприорецепторы Гольджи
- интрафузальные мышечные волокна или собственно проприорецепторы
- альфа – мотонейроны
- тормозные клетки Реншоу.
Рецепторы Гольджи возбуждаются при сокращении мышцы. Возбуждение идет к тормозным клеткам Реншоу в передних рогах серого вещества спинного мозга. Их возбуждение приводит к торможению альфа–мотонейронов и, следовательно, к расслаблению мышцы.
Проприорецепторы (интрафузальные мышечные волокна) возбуждаются при увеличении длины мышцы (при расслаблении, при растяжении).
Возбуждение от мышечных веретен идет к альфа-мотонейронам, что в итоге приводит к сокращению мышцы, повышению ее тонуса.
Эти два процесса чередуются с большой скоростью, поэтому мышца постоянно находится в определенном тоническом состоянии.
Для регуляции тонуса (не поддержания, саморегуляции, а изменения) необходимо изменение возбудимости проприорецепторов (интрафузальных волокон), что достигается с помощью гамма-мотонейронов.
Для регуляции мышечного тонуса (активного изменения тонуса, перераспределения его, обеспечения координированных движений) необходимо изменение чувствительности (повышение возбудимости, снижение порога чувствительности) проприорецепторов (интрафузальных мышечных волокон или мышечных веретен), что достигается с помощью гамма-мотонейронов. Гамма-мотонейроны вызывают сокращение мышечных отростков капсулы проприорецепторов, натяжение капсулы и, следовательно, повышается чувствительность проприорецепторов.
Гамма-мотонейроны находятся под влиянием вышерасположенных нервных центров продолговатого, среднего мозга и др.
Функции СМ:
1. Проводниковая (обеспечение связи в обоих направлениях);
2. Собственно рефлекторная (сегментарная).
Между ними сложные взаимоотношения: подчинение сегментарной деятельности надсегментарным центрам различных функциональных уровней.
СТВОЛ МОЗГА
Включает продолговатый мозг, мост, средний мозг, промежуточный мозг, мозжечок. В толще серого вещества продолговатого, моста, среднего, промежуточного мозга располагается ретикулярная формация.
Функции:
Организует рефлексы, обеспечивающие подготовку и реализацию различных форм поведения;
Осуществляет проводниковую функцию: через ствол мозга проходят в восходящем и нисходящем направлении пути, связывающие между собой структуры ЦНС;
Обеспечивает ассоциативную функцию, т.е. взаимодействие своих структур между собой, со спинным мозгом, базальными ганглиями и КБП.
Дата добавления: 2017-08-01; просмотров: 1033;